| 00:00 | 231 included nerve impulses and concentrating nervous or asteric gangling for the man |
|
| 00:15 | an f in the surrounding nerve fibers found that he could live muscle compaction |
|
| 00:21 | the battle. And the last shear were remain intact. So these |
|
| 00:32 | John and scientists quickly appreciated the significance young's final, but a advance was |
|
| 00:42 | actual amount that was fast enough to the techniques available at the time. |
|
| 00:47 | one of them designed for several hours isolated from people to be in a |
|
| 01:01 | to the discovery that sodium ions were concentrated outside the nerve cell and potassium |
|
| 01:07 | more concentrated inside. I recently, anti AON solution to the CP composition |
|
| 01:16 | were aided to theism of iron transport the ne on the five to be |
|
| 01:31 | through the C EM and into the ple. In these early techniques, |
|
| 01:42 | final glass chief is both consider the and secure with Clinton. It was |
|
| 02:09 | to introduce a high wire electric of the building between the inside and the |
|
| 02:14 | of the engine. But the formation the nerve impasse was slowly mounted. |
|
| 02:21 | detail study with any of the electrical devices that the late 1930 it wasn't |
|
| 02:28 | the 19 fifties following the war and of electronic equipment such as the capital |
|
| 02:33 | citizen that made the progress was But the cons is as a way |
|
| 02:43 | electrical potential that this not potential was by a trans movement of Syria is |
|
| 02:53 | comes to me, it started from square. This understanding then directed the |
|
| 03:07 | of drugs that no action potential So at local anesthetics, not usual |
|
| 03:14 | pain and devastated mind. Socially, knows the anesthetic, yeah, routinely |
|
| 03:39 | in anesthesia, lighter pain. But also talk about toxins. We'll talk |
|
| 03:49 | Tector the toxin. OK. So always, we can click on these |
|
| 03:53 | we watch them. Terrific. All still great. All these goldies. |
|
| 04:02 | So 100 years ago, 19 they could record or they could have |
|
| 04:09 | axons, they saw some something going with electrodes, but they couldn't |
|
| 04:14 | they could record slow changes in membrane , but they couldn't capture the action |
|
| 04:19 | until about 20 years later, the potential dynamics. And that's what I've |
|
| 04:25 | for you here. And this is to repeat and this is going to |
|
| 04:29 | again and then it's gonna repeat And then I have a whole diagram |
|
| 04:34 | you on action potential. So I in this folder also. Uh So |
|
| 04:39 | me exit out of this for a and show you what I mean by |
|
| 04:44 | ? Uh we go to the canvas oh maybe it's not here but I |
|
| 04:51 | uh I will make sure it's But in other words that these are |
|
| 04:58 | of the values you see that are here and these are the values that |
|
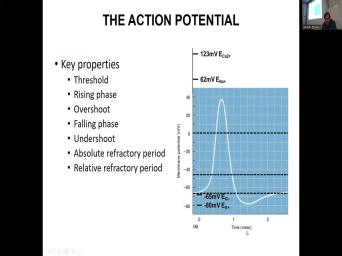
| 05:03 | gonna hold you responsible for the OK. Equilibrium potential for potassium equilibrium |
|
| 05:12 | for fluoride equilibrium potential for sodium equilibrium for calcium, the membrane potential VM |
|
| 05:22 | calculated using Goldman equation. Eken A equilibrium potentials are calculated using Nernst |
|
| 05:34 | The difference between GM. So this line and this action potential is the |
|
| 05:45 | membrane potential. So this is the value is this white line. This |
|
| 05:52 | resting membrane potential which is actually the to chloride and you'll say wait a |
|
| 05:58 | . But you just said something about leaking everywhere. It's influenced the most |
|
| 06:04 | potassium chloride channels are actually not open addressing me and potential. But the |
|
| 06:11 | value is the closest to to chloride . The other thing here this is |
|
| 06:19 | threshold for action potential. This dashed , this slide next slide or whatever |
|
| 06:27 | you choose are great slides, what called to mark your notes or mark |
|
| 06:33 | information. So for example, you put all of the information here rising |
|
| 06:39 | following phase we study in the new influx the E flux you know NAKDT |
|
| 06:46 | is being activated here. And uh we talked about the driving force and |
|
| 06:54 | driving force I explained to you the between the equilibrium potentials and the membrane |
|
| 07:01 | . So right here, for at the peak of the action |
|
| 07:06 | there is little difference between membrane potential equilibrium potential for sodium, there is |
|
| 07:13 | a difference of about 20 millivolts or , but it's small. So the |
|
| 07:18 | force here for sodium is small. the difference between when the membrane potential |
|
| 07:24 | at the peak and equilibrium for potential potassium is huge. Therefore, at |
|
| 07:33 | peak of the action potential, the force for potassium is much greater. |
|
| 07:39 | . That's an important component when we the action potential dynamics. Now |
|
| 07:44 | when the uh action uh when the membrane potential recovers the rustling memory potential |
|
| 07:52 | , you will say, well then driving force for potassium is not that |
|
| 07:57 | because there is not much difference between membrane potential and equilibrium potential for |
|
| 08:04 | But potassium is special and it has channels and it's still leaking. So |
|
| 08:10 | just nature. The threshold value rising overshoot, falling phase, undershoot during |
|
| 08:21 | action potential during this rising and falling right here, when the action potential |
|
| 08:28 | the threshold value and before it returns the threshold value, it's called an |
|
| 08:37 | refractory period. So if I were put a, put a box around |
|
| 08:45 | this area here, this would be absolute refractory period here that means that |
|
| 08:55 | this period, during this period, here, there cannot be another action |
|
| 09:04 | produced. And this right here when crosses back past the past the threshold |
|
| 09:16 | and before it rep polarizes in this , this is called the relative refractory |
|
| 09:23 | . So this area is a relative period. Because if there was a |
|
| 09:27 | enough stimulation during this time, there be a possibility that the plasma membrane |
|
| 09:34 | generate another action with it. So an aracy period, there can be |
|
| 09:40 | way for another actual potential to be . But it can be possible during |
|
| 09:45 | refractory period that is relative refractory OK. So the ups and downs |
|
| 09:54 | an action potential oscilloscope used to study potentials. Now there are digital |
|
| 10:01 | digital oteo copes the same things that talked about. Remember that these values |
|
| 10:08 | you're seeing here are the values that taken from the previous tables. When |
|
| 10:15 | talked about equilibrium potentials and N equation ring membrane potential and plug them into |
|
| 10:21 | scale here, you have to have number and depolarize past the threshold. |
|
| 10:28 | you depolarize the number and past the , the action potential is going to |
|
| 10:34 | because voltage gated sodium channels are going open a lot of them and very |
|
| 10:40 | and it will it is going to an all or none event. So |
|
| 10:44 | the numbering touches this threshold value, cannot go back down, it is |
|
| 10:50 | to produce this all or non it's called all or none because all |
|
| 10:54 | the action potentials will be always about same size and amplitude and about the |
|
| 11:00 | in duration. So about the similar and form that they will have. |
|
| 11:06 | this is all or none event, these depolarizations, some of these depolarizations |
|
| 11:12 | be small, some of them can large. Some of the depolarizations can |
|
| 11:16 | followed by hyper polarization. These are events. Uh So synaptic potentials are |
|
| 11:24 | typically graded potentials and action potential is or none. There's no gradation, |
|
| 11:32 | cannot have one action potential. 48 in amplitude and the next 1, |
|
| 11:37 | millivolts in amplitude will all be about millivolts or so and will fluctuate slightly |
|
| 11:44 | that particular change in the membrane So once the sodium starts entering into |
|
| 11:51 | cell, it opens up voltage gated channels right here and more sodium starts |
|
| 12:00 | into the cell. More sodium channels up. So these are gated by |
|
| 12:05 | again. So positive voltage will open channels. It will be flexing in |
|
| 12:10 | depolarization, more sodium multipolarization, more , more depolarization, more sodium, |
|
| 12:16 | depolarization, more sodium, it's called feedback loop. And what sodium is |
|
| 12:24 | is sodium is driving the VM which this white line, the membrane potential |
|
| 12:30 | the equilibrium potential for sodium. It all of my channels are open. |
|
| 12:36 | want to basically equalize these chemical and gradients or these forces. And I |
|
| 12:44 | to drive the membrane potential to my potential value. But it fails to |
|
| 12:49 | so because sodium channels also close very , they inactivate. And the second |
|
| 12:58 | is the closer the membrane potential is the equilibrium potential. The smaller is |
|
| 13:03 | driving force. Remember, the driving is the difference between, between VM |
|
| 13:09 | here and equilibrium potentials for given So now at this stage, the |
|
| 13:15 | channels close and there's small driving force sodium and potassium channels are all |
|
| 13:24 | So sodium is going in, in, in in and sodium channels |
|
| 13:29 | . It never reaches equilibrium potential. channels are now open and potassium now |
|
| 13:35 | the number and potential towards its own potential value because all of the channels |
|
| 13:42 | open and it almost suceeds succeeds to that when it goes through this um |
|
| 13:50 | and finally gets rebuild back to the potential value at r using the ionic |
|
| 13:57 | . So as you can see driving and the opening of the channel, |
|
| 14:02 | permeability for a specific ion are probably two most important things. So we'll |
|
| 14:06 | back and talk about it again on . Once again, I appreciate your |
|
| 14:11 | . Thank you for being here and sure you come the next couple of |
|
| 14:14 | . I'll be taking attendance. Thank |
|