| 00:02 | The second lecture on wiring the And we may even have a third |
|
| 00:09 | on it uh as we go through lot of interesting material in chapter 23 |
|
| 00:16 | particular, right. So the three of pathway formation we started discussing is |
|
| 00:29 | of all, there is pathway And again, we're using this uh |
|
| 00:33 | system example because you have the KS . And so the axons that are |
|
| 00:40 | , they have to decide if they're to go stay on the same side |
|
| 00:44 | the lateral or if they're gonna go uh fun the lateral on the other |
|
| 00:49 | . And then they have to decide they're going to innervate the medial genic |
|
| 00:54 | the lateral manic nucleus. And then have to decide which layer in the |
|
| 00:59 | genicular nucleus they're going to innovate. what happens during this growth? How |
|
| 01:08 | you have this kind of a allegation the of the axon as it is |
|
| 01:14 | ? Now, once you have a have dendrites, an important part is |
|
| 01:20 | only to uh be someplace but also your processes in such a way that |
|
| 01:28 | contact and you connect to the neighboring and neighboring sounds. And so the |
|
| 01:38 | this development process was happening when the are growing, the growing tip of |
|
| 01:48 | neuron, which is the axon really referred to as the growth cone. |
|
| 01:56 | what you can see is you have inside there and you have of course |
|
| 02:04 | and you also have lemo podia and smaller uh almost tentacle like structures called |
|
| 02:14 | Philippo. And essentially it is going act almost like an octopus at the |
|
| 02:21 | of the ocean, feeling things around it is extending for it to |
|
| 02:27 | It also has to have a uh of the underlying cyto skeletal elements, |
|
| 02:35 | the smaller ones that act them. now the other thing is you sometimes |
|
| 02:43 | several axons that interconnect and recognize each uh as axons with these cam molecules |
|
| 02:53 | cell in his and adhesion molecules. they recognize each other and they can |
|
| 03:01 | now kind of a group themselves in same fiber bundle and then potentially extend |
|
| 03:08 | the same direction and on the extracellular , uh there's an expression of these |
|
| 03:16 | molecules and on the axons, there's expression of these integra molecules. So |
|
| 03:23 | basically kind of a crawl along forming, they interact with other axons |
|
| 03:29 | then they sense the extracellular matrix environment interacting with that extracellular matrix. In |
|
| 03:39 | case, laminin with integrin and essentially their final sides of the connections. |
|
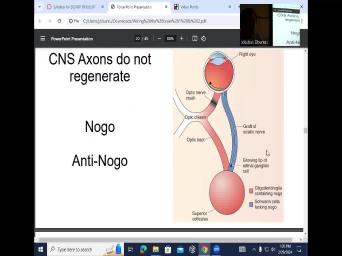
| 03:51 | it's a it's a really big question , in neuroscience, we don't make |
|
| 03:59 | lot of the stem cells that we about. Only 700 we cannot regenerate |
|
| 04:05 | . So if you have traumatic brain , you cannot regenerate those nes, |
|
| 04:12 | cannot regenerate axons that get cuts, example, during traumatic brain injuries, |
|
| 04:20 | you have regeneration, although sometimes it's depending on the type of the accident |
|
| 04:28 | the periphery. And the question is , what do you know that is |
|
| 04:38 | about peripheral nerves versus CNS nerves? thing that you know is that they |
|
| 04:46 | different myelination around them. Lugo Denver versus Schwan sal. But the other |
|
| 04:55 | is that they actually have a different . And that's really the the the |
|
| 05:07 | answer to this why PNS cells regenerate CNS cells do not regenerate. So |
|
| 05:20 | this case, what you have is have a graft of sciatic nerve and |
|
| 05:27 | graft of sciatic nerve, it has growing tip of retinal ganglion cell |
|
| 05:34 | So you have retinal ganglion cells. retinal ganglion cells would typically come out |
|
| 05:41 | the optic disc right here and form the right optic nerve on this |
|
| 05:49 | But optic nerve here has been it has been crushed. And instead |
|
| 05:57 | allowing for these retinal neurons that we projecting through the nerve to the co |
|
| 06:04 | the tract, you now lay a of sciatic nerve which is swan |
|
| 06:16 | which is peripheral nerve. Ok. what you have done is the red |
|
| 06:23 | the central nerves, the central the blue is you borrowed it from |
|
| 06:30 | periphery and he essentially like stuck it the right eye and said here, |
|
| 06:38 | a highway for this retinal cell now travel down and retinal cells. If |
|
| 06:45 | cut this, if you crush this , if you crush, cut |
|
| 06:50 | those retinal cells will not, that nerve will not regrow, they will |
|
| 06:56 | be able to travel. You their axons are not going to be |
|
| 07:01 | to reconnect again. So you provide graph and it says, OK, |
|
| 07:06 | use you and I'll regrow myself. this axon these axons have been cut |
|
| 07:13 | . OK? Maybe it's not uh clear, it was dive room. |
|
| 07:24 | so here is the retinal ganglion cells are putting their output through the optic |
|
| 07:32 | . OK. This is optic And then what happened is this, |
|
| 07:36 | is cut. So the axons of retinal ganglion cells, they're cut. |
|
| 07:43 | right. And they, they won't you'll for forever, not have any |
|
| 07:50 | uh going out of that eye. now you said, well, you |
|
| 07:59 | what? Here, there's a, a graph, there is a |
|
| 08:04 | you cannot go this way, you're , you can't regenerate. What if |
|
| 08:08 | put this peripheral Chuan highway and then of a sudden the south start migrating |
|
| 08:17 | and they're moving in this direction, in this direction moving in this direction |
|
| 08:23 | they encounter the area covered, it's , again covered by oligodendroglia. So |
|
| 08:35 | , there's no growth. So they of grow in to the, in |
|
| 08:40 | case, it's superior colliculus instead but it could be repeated with LGM |
|
| 08:49 | well. And that is oligodendroglia cells this no go molecule, obviously, |
|
| 09:00 | go is no go no regeneration. the Schwan sos are lacking no |
|
| 09:11 | And because of that, even the axon from the retina can now grow |
|
| 09:17 | not graft, but it cannot reinsert back into the, into the central |
|
| 09:23 | , central nervous system environment. What's ? I I have really poor |
|
| 09:38 | It doesn't have what effects they intended that. It's not really. Uh |
|
| 09:44 | , I mean, we would like have sciatic nerve grafts, but |
|
| 09:47 | that's the problem is that it can be tested experimentally. So the the |
|
| 09:55 | is always to try to regrow the nervous axons. Uh But if you |
|
| 10:04 | anti no go, if you inject , no go, you have anti |
|
| 10:10 | go antibodies, you can actually promote of that growth in the, in |
|
| 10:19 | , in the central, in the zones also. OK. So you |
|
| 10:24 | to block nogo. So that's an area uh for anything to do with |
|
| 10:32 | and grafting. So these axons, know, they are having interactions with |
|
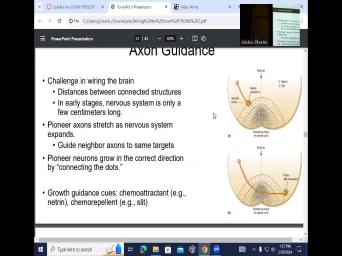
| 10:40 | other. They're having interactions with the environment, challenges and wiring brain or |
|
| 10:48 | between connected structures. Sometimes they can quite far from the eye, from |
|
| 10:55 | retina, the thalamus and the LGN then in the occipital lobe, |
|
| 11:00 | that's this axon has to travel quite in early stages. Nervous system is |
|
| 11:06 | a few centimeters long, but it becomes much longer. And so |
|
| 11:10 | think that the way this happens though kind of a like lema li |
|
| 11:14 | ali podia is feeling around like a . It's it's feeling around who it |
|
| 11:20 | associate with cell adhesion molecules where and it can associate with it extracellular |
|
| 11:28 | And the last part of it is there seems to be these pioneer axons |
|
| 11:34 | they are guided, guiding other neuro to the same targets. And it |
|
| 11:45 | to be that uh pioneer axons and growth of the pioneer axons, they're |
|
| 11:53 | kind of a paving the pathway let's say neuronal pathway where that axon |
|
| 12:03 | gonna be surrounded by 1015 other axons the way it is done and that's |
|
| 12:09 | it's called, it's connecting the So it's connecting this area of the |
|
| 12:14 | where it really needs to be in , the way this happens is again |
|
| 12:19 | that it happens that the growing growth are gonna contain natural receptors. And |
|
| 12:28 | have two, this is in the of the spinal cord. So ventral |
|
| 12:34 | uh dorsal uh surface of the spinal and then the ventral midline of the |
|
| 12:42 | cord got very high levels of expression me and slit. And they, |
|
| 12:50 | molecules are chemo attractants for neru chemo means that it's like, hey, |
|
| 12:57 | attracting that growth cone and chemo repellents , it's saying go away from |
|
| 13:03 | go away from here. And so growth cone, it has an expression |
|
| 13:08 | Nephron receptors. But also later, can see that it has an expression |
|
| 13:13 | robo or uh slit receptors. So going on here? First of |
|
| 13:20 | there is a differential extraction of these and slit molecules, but it is |
|
| 13:28 | really strong attraction because it contains Merin . If there's growing stage, a |
|
| 13:34 | more Merin receptors are expresses, trying get, be attracted into the midline |
|
| 13:41 | . But then gradually which happens during development, there might be different ratios |
|
| 13:48 | changes of these receptors. And a bit later, there might be a |
|
| 13:53 | expression of provo slip suckers. If example, you have a neuron that |
|
| 14:03 | has a lot of Nephron receptor and take Losito and glib one. |
|
| 14:09 | if this neuron also now starts expressing a different neuron that expresses ro slipp |
|
| 14:17 | or the same one that expresses in la later stage is now going to |
|
| 14:22 | attracted to areas that are more dominated slit. So it's kind of more |
|
| 14:28 | both what the growth to expresses and what it is in this chemo attracting |
|
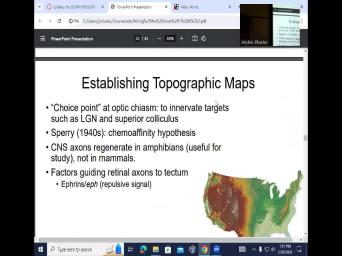
| 14:35 | chemo repellent soup as they call uh establishing topographic maps, optics as a |
|
| 14:48 | point and optics to the targets such LGM and superior Colliculus. So the |
|
| 14:54 | split lateral jar nucleus uh is not only target, not the retinal output |
|
| 15:01 | as we saw just recently the superior . So there's this chemo affinity hypothesis |
|
| 15:08 | was developed in the forties. And other interesting thing is that there are |
|
| 15:16 | animals and in particular amphibians that are useful in studies of regeneration. So |
|
| 15:26 | know that if you like cut off tail and it's actually with their spinal |
|
| 15:32 | , they will, they will So the uh these amphibians can regrow |
|
| 15:38 | whole arm uh or uh maybe they're arms and amphibians but they uh |
|
| 15:49 | their limbs but not in, not mammals. Uh So what we |
|
| 15:55 | what we learn in amphibians could be to humans, potentially. I |
|
| 16:02 | these are some of the biggest questions think that the humanity is trying to |
|
| 16:07 | . We, we've been able, think in the last 100 couple of |
|
| 16:11 | years, maybe not 100 maybe a 100 years to double our lifespan. |
|
| 16:17 | you, we used to live just to the thirties, forties. |
|
| 16:23 | the life expectancy in the seventies depending , on many different things and the |
|
| 16:30 | here in and stuff like that. , but so can we live |
|
| 16:37 | And what would it take for us live longer? What would it take |
|
| 16:41 | us to contain, having enough of brain cells to be conscious that we're |
|
| 16:47 | , not just the muscles living you know, and breathing. And |
|
| 16:52 | you're starting to think about, what are the environments and these other |
|
| 16:57 | and models that allow for axons to , for limbs, to regrow, |
|
| 17:03 | tails and spinal cords to regrow. we harvest something from their chemical soup |
|
| 17:12 | influence the capabilities of the regeneration and mammalian brains. And it's a long |
|
| 17:21 | , but I think that we learn lot from am amphibians, we can |
|
| 17:26 | it to mammals. But eventually, still need to apply it to mammals |
|
| 17:31 | we understand what it does to humans mammal models, of course, uh |
|
| 17:40 | guiding retinal axons to tectum. So it's eeph uh producing a repulsive signal |
|
| 17:52 | tectum is also superior colliculus. Uh tectum versus tantum, tectum is superior |
|
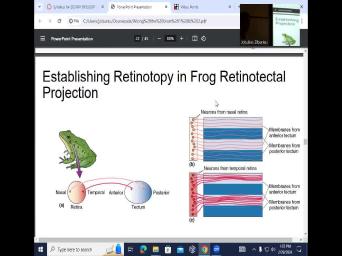
| 18:03 | . This is really pretty cool because have the retina, nasal, temporal |
|
| 18:11 | , anterior, posterior. So in of these primitive animals, tectum actually |
|
| 18:18 | lateral geniculate nucleus. OK. And of the primitive animals, when you |
|
| 18:25 | about tectum and amphibians, this is equivalent of the thalamus and the lateral |
|
| 18:31 | . No, that is the main processing center of the visual information. |
|
| 18:39 | so here you have membranes that have laid from anterior attack them and the |
|
| 18:46 | that have been laid from posterior tectum neurons that are taken from the nasal |
|
| 18:53 | . So you take the nasal retina , you take neurons on the nasal |
|
| 18:58 | , you harvest them, you put in these plates now and you provide |
|
| 19:04 | the highways to grow and you provided membranes from anterior and posterior, anterior |
|
| 19:11 | posterior and nasal neurons are going to able to grow in develops. So |
|
| 19:20 | , remember what happens with with nasal this eye, they can cross over |
|
| 19:27 | what happens with temporal, they stay the same side. And now you |
|
| 19:34 | neurons from this other side right from the temporal retina and you lay |
|
| 19:41 | out and lay the same highway. they're like nope, we're all here |
|
| 19:44 | go to the anterior tact with only the anterior tact him. So this |
|
| 19:54 | a simple example of the anterior posterior who goes where. But it also |
|
| 20:03 | applicable to essentially what we understand and . And obviously, we don't understand |
|
| 20:10 | kind of a we haven't been able isolate fresh human retinal cells and lay |
|
| 20:17 | the graft. So a lot of we know is still at the lower |
|
| 20:21 | animals and studies and and and animals froggies, neuromuscular junction and neuromuscular |
|
| 20:33 | So you have a growing motor neuron this is a growing motor neuron and |
|
| 20:40 | is secreting the agri molecules. So the same as in C MS, |
|
| 20:47 | need to find each other and form in the P MS, the axons |
|
| 20:53 | out to contact the muscles, they need to form synapses. And it |
|
| 20:59 | here that you have these ach receptors are sort of a migrating. |
|
| 21:07 | it's a fluid mosaic model in the membrane. And that means that they're |
|
| 21:12 | really sequestered and uh kind of a up in one particular area. So |
|
| 21:21 | out uh outgrowth of these axonal codes the grin molecules. The grin molecules |
|
| 21:31 | interact with the agri receptor which is in this interaction will lead through this |
|
| 21:41 | and molecule will lead to the clustering ach receptors. And once these ach |
|
| 21:49 | get clustered in the postsynaptic membrane, now have the ability to form a |
|
| 21:59 | . And now these ach receptors are just like migrating and floating everywhere |
|
| 22:05 | but instead they get like sequestered in area which is now going to become |
|
| 22:11 | junction. So on the postsynaptic it's not only on the presynaptic side |
|
| 22:18 | also what is happening postsynaptic and how have to match the presynaptic forming side |
|
| 22:26 | the active zone with a postsynaptic and postsynaptic uh densities on dendrites are here |
|
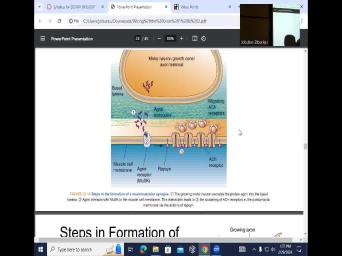
| 22:35 | the neuromuscular junction on the muscle, steps of the formation of the |
|
| 22:45 | So you have the dendritic to the that contacts the axon HM. And |
|
| 22:53 | you have the synoptic vesicle formation and presynaptic active zone formation. And then |
|
| 23:01 | have the clustering of these foam receptors are spatially going to be juxtaposed to |
|
| 23:09 | um synaptic vesicles and the axons. we are born with a lot more |
|
| 23:20 | and a lot more neurons than we up in adult brain. This is |
|
| 23:25 | process of synoptic city that will continue uh uh for the next I think |
|
| 23:34 | lectures or so in in in So what happens is during the development |
|
| 23:46 | you first have a lot of inputs a lot of them are connected to |
|
| 23:57 | and a lot of them are actually in non-specific ways to almost like everything |
|
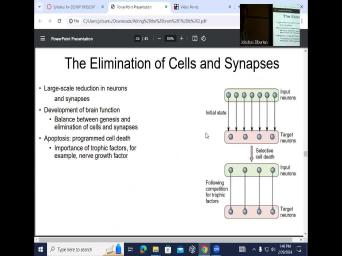
| 24:03 | connected to everything. And then there uh elimination of some of these not |
|
| 24:14 | synopsis but also nerves. So we end up eliminating cells, we're born |
|
| 24:23 | more cells and we end up as . Therefore, we also have more |
|
| 24:27 | . So we end up with adults . So how do you go through |
|
| 24:32 | process of what we call selected cell ? And in this case, when |
|
| 24:37 | talk about cell death, we're talking apoptosis, which is programmed cell |
|
| 24:45 | And it's different from necrosis. Necrosis neuronal cell death that gets induced due |
|
| 24:52 | injury due to infection, due to inflammation and things that we uh touched |
|
| 24:59 | uh a little bit in this course . So there is a fine balance |
|
| 25:08 | the birth of new cells and synopsis elimination if you eliminate too many cells |
|
| 25:16 | too many synopsis. It's not If you leave some of these connections |
|
| 25:22 | are unrefined, it can potentially lead conditions like autism spectrum disorders. Because |
|
| 25:32 | of the autism theories is that there a impairment and pruning in certain regions |
|
| 25:39 | the brain of these synapses and interconnectivity it is pretty highly inheritable condition. |
|
| 25:50 | what is the trophic factors? And first one is NGF or nerve growth |
|
| 25:58 | and there's many of these growth factors BDNF, brain derived nerve factor, |
|
| 26:05 | nerve growth factor. So, there these trophic factors that are present during |
|
| 26:13 | development and wiring and these trophic NG FBD NF to have their own |
|
| 26:22 | . So they're typically truck receptors, in kinase receptors and these tropic factors |
|
| 26:29 | not there in huge abundance and they're there forever. So there's during the |
|
| 26:37 | development, there's increased production of the factors and they're necessary really to control |
|
| 26:44 | cell survival and the con activity between input neurons and certain target neurons. |
|
| 26:52 | also they're very much involved in essentially apoptosis, which is regulating the |
|
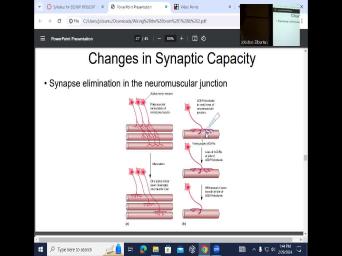
| 27:05 | So that so back to neuromuscular synapse elimination in the neuromuscular junction. |
|
| 27:21 | what happens in neuromuscular junction is you can see the south motor neuron |
|
| 27:29 | it will innervate 123 muscle fibers. we call this poly neuronal innervation of |
|
| 27:38 | muscle. But when we look in mature muscles during maturation process, we |
|
| 27:45 | only a single alpha motor neuron innervating single motor muscle fiber. So this |
|
| 27:54 | been refined, we call them. has been anatomically segregated into specific fibers |
|
| 28:03 | anatomically refined for specific connectivity. Uh happens if during this process, you |
|
| 28:13 | acetylcholine receptor. So, acetylcholine receptor can be accomplished with acetylcholine antagonist receptor |
|
| 28:24 | . And you know, one of , it's called curare. So if |
|
| 28:28 | apply curare on this neuromuscular junction block , you have loss of ach receptors |
|
| 28:39 | the side of the ach receptor And guess what happens to the Saxon |
|
| 28:46 | axon is no longer interested in staying that area. So once again, |
|
| 28:52 | not only dependent on how the Saxon fueling things up, but it's also |
|
| 28:59 | on the response it's gonna get from muscle. And if this, there's |
|
| 29:04 | response from stimulating this area of the , because they see the colon receptors |
|
| 29:09 | blocked. Therefore, there's no action . Therefore, it's the muscle is |
|
| 29:16 | improperly. So it's just going to , ok, never mind, I'm |
|
| 29:21 | just stay in the area where the receptors are active. So you have |
|
| 29:27 | presynaptic and postsynaptic activity dependent interactions that , that will drive refinement. And |
|
| 29:37 | in general, when we talk about plasticity or refinement of the synopsis, |
|
| 29:42 | we have finite amount of tissue in brains, we have finite amount of |
|
| 29:48 | in our adult brains and then we finite amount of synopsis, we can |
|
| 29:54 | in our brains. But what we do is we can increase the capacity |
|
| 29:59 | increase the specificity and the communication between parts of the brain. So this |
|
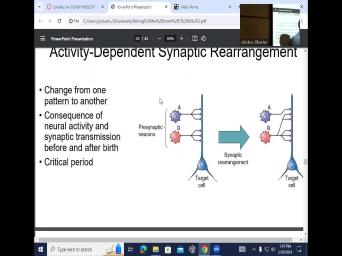
| 30:10 | some of the cartoonish representations of what during activity dependent synoptic rearrangement. And |
|
| 30:19 | keep talking about that today. The lecture also is when you have a |
|
| 30:25 | from one pattern to another. So you have a B two neurons, |
|
| 30:31 | one of them have three axonal collaterals innovate the target parameter C A and |
|
| 30:38 | have three. And what if he extremely active? And A is |
|
| 30:50 | And this is what happens uh right when, before birth and right after |
|
| 30:59 | is when we have neural activity. why, why do I say |
|
| 31:04 | Wait a second neural activity? we didn't have any neural activity until |
|
| 31:09 | . We were talking about the growth , fill out this area, the |
|
| 31:16 | filling out this area. And then just started talking about how you then |
|
| 31:21 | to have postsynaptic receptors come in on other side. And now we finally |
|
| 31:27 | the activity because we have the presynaptic , we have the postsynaptic receptors now |
|
| 31:35 | in the synopsis. So now we activity and so it happens that cell |
|
| 31:40 | is extremely active and cell A is . And so during this process, |
|
| 31:45 | happens is cell B has a much stronger capacity to influence this target |
|
| 31:54 | not capacity in the electrical terms, to influence the target itself. |
|
| 31:59 | it has five synopses, it's really stronger compared to this guy and this |
|
| 32:05 | continues not firing, may lose his synapse. And if you lose those |
|
| 32:10 | synapse, you may not even exist . And so this synaptic rearrangement and |
|
| 32:20 | refinement in the Mussels in the CNS is happening during what we call a |
|
| 32:27 | period of development. It doesn't last . Even in uh uh last |
|
| 32:36 | we talked about the fact that there's many more stem cells during this |
|
| 32:41 | I mean, they're like at some of gestation, they're just all stem |
|
| 32:47 | , you know, and then we're to 700 a day. And so |
|
| 32:57 | the time when the brain is forming , where it's, it's could be |
|
| 33:03 | . I think we could be losing of thousands of neurons a day through |
|
| 33:08 | process of rearrangement refinement and uh program death and this process is activity dependent |
|
| 33:17 | and this critical period doesn't last So it's special and it's related to |
|
| 33:24 | very early activation of the synapses, migration, but this very early activation |
|
| 33:33 | the synopsis where that what I call soup from which neurons were differentiating and |
|
| 33:42 | in different locations. This primordial soup still really favorable to change things to |
|
| 33:51 | , to refine things in the but it doesn't last for long. |
|
| 33:59 | let's talk about segregation of LGN inputs the stride cortex, which is shown |
|
| 34:05 | the slide. But before we do , I'm gonna tell you a little |
|
| 34:10 | about what I did for my phd . And I'm gonna give you a |
|
| 34:15 | talk. What the chop talk I don't have chalk but I have |
|
| 34:22 | . So talk talk is when you something and you do your best |
|
| 34:27 | to draw of the. So when was studying is I was studying |
|
| 34:40 | That was my model. I also studying mice, but this was my |
|
| 34:49 | mammalian rodent model. The time period was looking at, I was looking |
|
| 34:58 | P zero to P 21. Thank . This is the time P stands |
|
| 35:07 | postnatal. So I was studying these animals. The area that I was |
|
| 35:16 | in is retinal, geniculate athletes rest love. OK. From retina and |
|
| 35:39 | the lateral geniculate influence. My rodent was in vitro. Mhm which means |
|
| 35:55 | I wasn't studying activity in the whole or the brain, but we were |
|
| 36:01 | activity in a dish. However, our situation, it was kind of |
|
| 36:11 | and it could almost be called ex , not in vivo inside but ex |
|
| 36:21 | outside. And the reason for it that as I was doing my |
|
| 36:31 | if you recall, we have this here, we have two eyes, |
|
| 36:36 | have projections that cross over and actually have the same and the and the |
|
| 36:45 | the rodents. So you also have projections that stay on the same |
|
| 36:53 | Oh Wait a second. So the the cross is over, crosses over |
|
| 37:01 | on the same side. So when entered the lab, this is the |
|
| 37:10 | year, these are the fibers going the LGM. When I entered into |
|
| 37:20 | , into the lab, we had preparation where LGM was cut out. |
|
| 37:27 | is here is optic trapped. And we were stimulating, my mentors were |
|
| 37:37 | my great mentor in electrophysiology was a professor, Doctor Fu San Lu. |
|
| 37:45 | And uh we would stimulate optic tract then we record from these neurons in |
|
| 37:53 | LGM with in intercellular recording electrons. so we would shock the track here |
|
| 38:03 | this track has been cut. And one of the issues was that |
|
| 38:11 | it's problematic because when you apply a on the optic tract, you are |
|
| 38:19 | stimulating blue and black and blue and fibers that are intermixed. Uh So |
|
| 38:32 | I came into the lab, I a liking to doing dissections of their |
|
| 38:38 | . And I said, let me with this a little bit, figure |
|
| 38:41 | out. And my dissertation then became this is X vivo because in really |
|
| 38:49 | animals, really small rats, this is really small, it's not like |
|
| 38:56 | slice, but it's like one giant and that's why it's ex vivo. |
|
| 39:02 | so when I looked at this What I was able to do is |
|
| 39:07 | was able to isolate both nerves. one LGM. Now I had the |
|
| 39:24 | of stimulating either either the blue fibers the black fiber. So it was |
|
| 39:36 | clear now that I was either stimulating ipsilateral inputs because I was stimulating |
|
| 39:45 | I was stimulating at the level of optic nerve. When you stimulate optic |
|
| 39:55 | on the left side, you know on the left LGM, you're only |
|
| 40:02 | to have ipsilateral input. When you this fiber optic nerve fiber here and |
|
| 40:12 | have the opposite side. LGM. know the only recordings you're gonna |
|
| 40:17 | if you stimulate this nerve, it's be contralateral. And this is what's |
|
| 40:24 | interesting about LGM LGM. At if you looked at it and you |
|
| 40:35 | contralateral versus ipsilateral, so you can the dice pumps. So later, |
|
| 40:44 | , you can inject the dye in eye and then you can inject |
|
| 40:50 | the dye into another eye. Let's that. So we're gonna have |
|
| 41:04 | oh We have this and we're gonna at one LGN here and we're gonna |
|
| 41:14 | at this LGN as a function of . So we're gonna inject green dye |
|
| 41:23 | to this side and inject black dye the side and we're gonna take LGM |
|
| 41:30 | just one side, right? And see where there is a distribution of |
|
| 41:40 | C inputs. These are IPs these are IPs inputs. Versus the |
|
| 41:49 | of the south and the inputs going that are contra the the other, |
|
| 41:58 | other side of Belgium was cut, all over. That means there's really |
|
| 42:06 | much specificity immediately after birth. The important thing to note is and |
|
| 42:17 | eyes open about P 14 post the 14. So they're more with their |
|
| 42:28 | uh closed at first. What happens we go to P seven? And |
|
| 42:37 | look at the LGM, remember the is contra we start seeing sure kind |
|
| 42:59 | a formation of two separate zones So you have the segregation, you |
|
| 43:06 | the refinement of these that are interspersed into what we call ipsilateral and contralateral |
|
| 43:18 | . Then finally, when you look P 21 find that OK, is |
|
| 43:40 | very clear IPY zone that is surrounded a dominant contralateral zone? And this |
|
| 43:50 | sort of what the adult inputs into lateral because that new look like. |
|
| 44:00 | look at the physiology during this time . There's some very interesting things that |
|
| 44:08 | going on. First of all, before the eyes open in these |
|
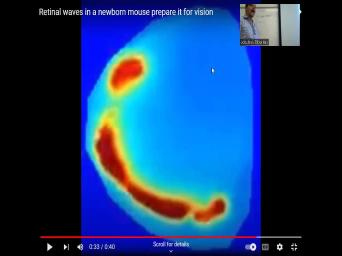
| 44:14 | I think I have this a little later before the eyes open in these |
|
| 44:21 | , the retinas of producing these spontaneous of activity. So the photoreceptors, |
|
| 44:29 | waves of activity are present in animals the photoreceptors are even functional. This |
|
| 44:36 | a code, there is an ongoing of repetitive activity inside the retina that |
|
| 44:44 | activity. And that code is constantly to find its way and connectivity into |
|
| 44:52 | lateral geniculate nucleus. If during early , we block these retinal waves, |
|
| 44:59 | do not get the proper wiring from retina to the LGM. We destroy |
|
| 45:05 | genicular connectivity. So these waves are important and they're happening spontaneously, which |
|
| 45:13 | that there's no stimulus. And in , they're happening before photo receptors are |
|
| 45:19 | . It means there's no photo transduction is going on. They're not being |
|
| 45:25 | by external stimuli. This code and waves are not unique in early development |
|
| 45:32 | neurons, just the rat up. lot of times, we have these |
|
| 45:40 | circuits that are very important in developing spinal cord circuits. These waves are |
|
| 45:49 | turtles, turtle retina and roden, uh frog re uh is it in |
|
| 46:00 | ? Well, we do so it's also present in mammals, but |
|
| 46:06 | have to go into into earlier So they are present in different parts |
|
| 46:16 | the body and they present sort of almost like a signal activity signal that |
|
| 46:24 | that is there that is going to partly how this is going to shape |
|
| 46:30 | intrinsic mechanisms, intrinsic physiology. when the eyes open at P |
|
| 46:40 | the final refinement is happening with the of the activity with the external |
|
| 46:46 | extrinsic stimulant, right? This play did I close that by accident pause |
|
| 47:25 | recording? I somehow missing my my uh it just gotten hidden from |
|
| 47:56 | So that is really, really uh from the anatomical. OK. So |
|
| 48:05 | I described to you is anatomical We haven't talked about activity, started |
|
| 48:12 | about activity. I said, let's at these really interesting retinal weights, |
|
| 48:18 | then there's early activity and we were to know what happens during early |
|
| 48:26 | And this is LGM. This is single song that LGM actually let me |
|
| 48:37 | a different, this is a So, and the LGM FP |
|
| 48:50 | this is a single cell and they it P 21. So when we |
|
| 48:58 | able to stimulate the tracks and the , we were able to determine how |
|
| 49:05 | synopses because we can do minimal UPSD . We were able to determine how |
|
| 49:11 | synopses each one of these, they're relay cells in the LGM. How |
|
| 49:18 | how many synapses these LGM relay cells receiving. And what we saw at |
|
| 49:26 | beginning is first, they were receiving lot of inputs and those inputs were |
|
| 49:39 | if so lateral and consol lateral. at T 21 the south um retina |
|
| 49:55 | LGM. This is retina to LGM one retinal ganglion cell to one relay |
|
| 50:07 | piano and just either contralateral or just what does that tell you? It |
|
| 50:15 | you that during early development. we talked about the visual system, |
|
| 50:19 | said like it's all monocular, it's monocular segregated LG monocular is segregated even |
|
| 50:24 | the layer of the cortex. And it only starts being binocular there 23 |
|
| 50:29 | the cortex. In fact, in early development, because of this mass |
|
| 50:35 | anatomical projections and connectivity to cells, have binocular cells. So early on |
|
| 50:43 | relay cells are receiving input from both because all of the anatomy there is |
|
| 50:51 | and they have a lot of And what happens you have this process |
|
| 50:57 | pruning. You have this process of and only the active synopsis, the |
|
| 51:05 | falls from the retina link up to one and it becomes 1 to 1 |
|
| 51:12 | . And that's where we start developing retinal topic map, one retinal gang |
|
| 51:18 | to one LGM cell. But then we get to cortex LGM cells will |
|
| 51:23 | of cortical cells. OK. So is all essentially uh about five years |
|
| 51:36 | my work only about uh a half it. Because when we talk about |
|
| 51:42 | plasticity, I'll tell you what else in the system as it concerns synaptic |
|
| 51:50 | also. But before I do let's move up a little bit into |
|
| 51:56 | cortex on the LGM. And we a lot of you have seen the |
|
| 52:03 | already. But essentially what this experiment is the following. This experiment is |
|
| 52:12 | short term deprivation experiment where an eyelid being sutured for a period of three |
|
| 52:21 | . As you recall, we have dominance and in the in the visual |
|
| 52:29 | . So you have eyelid, sutured three days and then you let animal |
|
| 52:34 | for a month and then you expose eyes to light. The one that |
|
| 52:39 | sutured and the one that was and you're recording responses in the |
|
| 52:44 | And after three days, there is shift toward more responsibility to the eye |
|
| 52:50 | was left open. If you repeat same experiment, this is during the |
|
| 52:56 | year, during sorry during the first of life. And why is that |
|
| 53:00 | ? Because here we're all looking at first month of life. P ZERO |
|
| 53:05 | only want is three weeks, another is one month of life. So |
|
| 53:10 | nearing the end of the biggest refinement . The biggest refinement period is happening |
|
| 53:18 | that P zero to P 21. we're coming out toward the tail |
|
| 53:24 | That means things are not gonna be plastic anymore that tail end, but |
|
| 53:29 | still the ability to kind of rearrange synoptic inputs. But if you close |
|
| 53:36 | that period, tail end period, almost entire week, six days, |
|
| 53:41 | eyelet, then once you reopen it month later, cortex doesn't care anymore |
|
| 53:47 | the eye that was closed. So have massive cortical rearrangement. If this |
|
| 53:51 | a normal eye, even after short deprivation, these are the LGM axons |
|
| 53:57 | into the cortex really bushy, very and robust. And the deprived eye |
|
| 54:03 | are now have been essentially altered and are far and few in between. |
|
| 54:09 | this is an activity dependent process. activity dependent, it's activity dependent |
|
| 54:14 | just like we're seeing here before eyes , after eyes open and even before |
|
| 54:21 | . So, plasticity during critical period development, if we look at how |
|
| 54:29 | can change this ocular dominance in the visual cortex, uh you can see |
|
| 54:38 | you can do these shifts and in , this is age in weeks uh |
|
| 54:45 | there is no really shift in ocular in the sense that we have about |
|
| 54:53 | weeks of from 100 to 0. it's from 0 to 100 right? |
|
| 55:00 | almost from 100 to 0 in six , 16 weeks. This is uh |
|
| 55:07 | time range for for human critical period in just uh the early life and |
|
| 55:17 | adolescence also. And then there's a of, there's a lot of pruning |
|
| 55:22 | going on in early adolescence. Your says that's why it's such a trying |
|
| 55:28 | to be a teenager. You modulatory influences. You guys remember the |
|
| 55:39 | systems retinal activity before birth. I into that a little bit earlier. |
|
| 55:47 | And then you have visual environment after . So the stimuli are different and |
|
| 55:53 | you have some enabling factors that are . So the chemicals, not just |
|
| 55:58 | receptors and those chemo repellents and chemo are going to be changing. Uh |
|
| 56:06 | the signaling and MD A is going be changing in the Synopsis Gaba recall |
|
| 56:12 | can be excited to in early developing good. And uh this is the |
|
| 56:21 | that we already discussed. So you locust Aurelius projections, uh and basal |
|
| 56:29 | complex projections. So you have anybody what those are. Locus Aurelius, |
|
| 56:42 | and basal chlorine easy. It's only that is there acetylcholine. So mono |
|
| 56:54 | in in any case, and this an experiment where you have normal input |
|
| 57:03 | motor immune systems and monocular deprivation And you have a shift in ocular |
|
| 57:09 | like you would expect. And now actually eliminate uh knife cuts right |
|
| 57:17 | You you, you literally cut the from the supply of norepinephrine and the |
|
| 57:26 | of acetylcholine. And now all of sudden there is no change in ocular |
|
| 57:34 | . You see that this is a in ocular dominance. If you sutured |
|
| 57:40 | eyelid, you would see the shift like you saw in these experiments. |
|
| 57:45 | is a change in ocular dominance. . There's a significant shift toward one |
|
| 57:53 | . There's a significant shift here toward side after the monocular deprivation because you |
|
| 58:00 | mono means. But now you cut on a mean input and you don't |
|
| 58:07 | the same anatomical rearrangement. So that you that these modulatory inputs play a |
|
| 58:15 | in the axonal refinement in this activity processes also. And we'll see that |
|
| 58:24 | uh over next lecture. Um So think we'll probably spend uh maybe quite |
|
| 58:33 | bit of time again in the next . Uh finishing things up, I |
|
| 58:39 | one order uh slide outside the But before we finish today, when |
|
| 58:44 | come back, next time, we continue talking about kind of a cellular |
|
| 58:49 | of plasticity, we'll go back to same systems, the retina geniculate |
|
| 58:56 | And we'll discuss at least two the rate code and the spy timing |
|
| 59:01 | on plasticity code by which neurons strengthen weaken their synapses. So that's gonna |
|
| 59:08 | next lecture. And I'll tell you some of my work that I did |
|
| 59:14 | I was a post doc. Uh before we end today, why do |
|
| 59:21 | periods? And wouldn't it be beautiful we could extend the critical period so |
|
| 59:29 | you would have the ability to you would have greater learning capacity so |
|
| 59:36 | maybe you could live longer, not dementia, not get depression. Three |
|
| 59:46 | that your book discusses why this plasticity as the axonal growth cone CSS. |
|
| 59:55 | it's no longer growth cone. Once contacts the posy side and has a |
|
| 60:02 | , one synaptic transmission matures. So another way of looking at the end |
|
| 60:10 | the critical period. And when cortical is constrained because you have significant anatomical |
|
| 60:19 | and development of these inhibitory circuits. it's not very easy for the brain |
|
| 60:24 | to be activated. And intrinsic inhibitory are actually late to, to |
|
| 60:32 | So that tells you that at the brain is very highly excitable and |
|
| 60:39 | later with the incoming inhibition, you have essentially restraint of that activity. |
|
| 60:50 | , developmental regulation of plasticity may help from CNS damage. So all of |
|
| 60:56 | mechanisms, not only the guidance but regrowth and the plasticity activity depend on |
|
| 61:06 | . We need to find all of different ways right from maybe sensory stimulations |
|
| 61:12 | activate the fibers in a certain Pharmaceutical drugs, uh chemo repellents, |
|
| 61:18 | , grafts that could be placed in for these cells to regrow in the |
|
| 61:25 | . All right. So we'll end today and we'll finish on really synaptic |
|
| 61:33 | . Although uh chapter 25 is more synaptic plasticity, but also more on |
|
| 61:38 | molecular kind of a level. Uh I will try to save and |
|
| 61:45 | Uh This is now, I think had two lectures. So I'll upload |
|
| 61:49 | two lectures and I will upload the readings on spike timing, dependent |
|
| 61:56 | And also what I've described here for . Uh because the, the chalk |
|
| 62:01 | elements will also be uh questions on quizzes and your, and your |
|
| 62:07 | OK. All right. Thank |
|