| 00:02 | OK. So we continue talking about membrane potential and we started talking about |
|
| 00:07 | pro channels because the ions that surround membrane from either the external side or |
|
| 00:16 | side, they cannot cross through the membrane. Ok. So what we |
|
| 00:21 | to do is we need to build channels. These channels when we talk |
|
| 00:27 | especially the action potential are gated by and they're specific or they're selected to |
|
| 00:34 | ions. We will also later in course, talk about light gated channels |
|
| 00:40 | we talk about receptor channels, uh regard to neural transmission and then mechanically |
|
| 00:46 | channels and these channels, if you all of these proteins are built out |
|
| 00:52 | these building blocks, and some of amino acids will have amino acid residues |
|
| 01:00 | the internal lining of these channels that be interacting with the ions. |
|
| 01:09 | ion pumps work differently. You'll notice channels are gated by voltage as we |
|
| 01:17 | . And by other things, the are driven by a TP. So |
|
| 01:22 | another reason why the brain consumes a of energy. It will need to |
|
| 01:27 | these pumps and these pumps will use lot of A TP and they will |
|
| 01:33 | ions against their concentration greatness. So will learn that these proteins will conduct |
|
| 01:41 | to that extracellular fluid where there is concentration of sodium. So against concentration |
|
| 01:48 | and this is going to pump in the same time, two potassium ions |
|
| 01:54 | intercellular which is against the concentration gradient potassium. And then of course, |
|
| 02:03 | is different pumps. Apart from just A T DNA K pumps that we |
|
| 02:09 | about, there are calcium pumps that transport calcium, there are different transporters |
|
| 02:17 | will also transport neurotransmitters. And if just imagine a a simple situation of |
|
| 02:27 | concentration chemistry as is illustrated here and have sodium chloride, a lot of |
|
| 02:35 | from this side of the membrane and no channels here. So sodium fluoride |
|
| 02:40 | pass through. If you insert channels the specific sodium fluoride, these two |
|
| 02:46 | are going to flow down to This is called infusion its chemical gradient |
|
| 02:52 | will be driven by chemical gradient onto side until there is an equal molar |
|
| 02:58 | equal concentration of these ions on each of the membrane. So note |
|
| 03:07 | that channels will allow crimes to be on concentration gradients. But that is |
|
| 03:14 | the only determinant as you learn, for voltage gated channels, they're permeable |
|
| 03:21 | specific ions and that there is a gradient or uneven distribution of charge across |
|
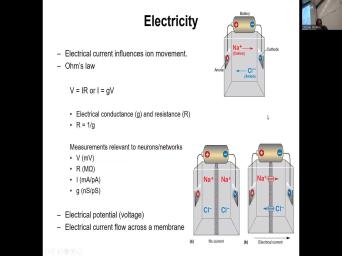
| 03:30 | plasma membrane. Now, some of basics of electricity that you may recall |
|
| 03:38 | high school or in college. Here review also is that for example, |
|
| 03:44 | have a battery which has the positive the negative and the positive is the |
|
| 03:50 | node and anions are negatively charged ions gonna be attracted to the positive end |
|
| 03:56 | the anade battery. The chode which negatively charged and of the battery will |
|
| 04:03 | attracting sodium ions to the negative sodium positively charged. So that tells |
|
| 04:11 | that apart from the chemical gradient and pure concentration dependent drive of ions they |
|
| 04:20 | charged. Therefore, there's going to an interaction and the flux of ions |
|
| 04:26 | not just going to depend purely on chemical gradient, but it's also going |
|
| 04:31 | depend on the electrical charge of these and the batteries which essentially form across |
|
| 04:41 | plasma membrane. So electrical currents will only influence ionic movement, it is |
|
| 04:48 | ionic movement that is the electrical And if you call arms law V |
|
| 04:56 | ir so V stands for voltage and neurons, the relevant scales that we're |
|
| 05:05 | in voltage is going to be mostly nodes so that we can pick up |
|
| 05:10 | activity as well. Uh I stands current and the neuro relevant scale is |
|
| 05:19 | many anthers or people anthers and r for resistance, which is measured in |
|
| 05:30 | and the relevant scales for neurons as and mega os. And that is |
|
| 05:38 | neurons at very small cells, about micrometers in diameter. The smaller the |
|
| 05:45 | , the higher the resistance. electrical conductance is the inverse of the |
|
| 05:54 | . So you can rewrite R is one over G. So you can |
|
| 06:00 | S Law B equals ir or I equal G conductance times the change in |
|
| 06:13 | and conductance. Uh It relevant scales measurements for neurons is in nano semen |
|
| 06:22 | pick a semen. So electrical the potential of the battery is |
|
| 06:28 | And when you put a volt meter across the two ends of the battery |
|
| 06:34 | in your house AA or AAA, says 1.5 volts. So you this |
|
| 06:40 | the the voltage OK. And that's there is an equal separation of charge |
|
| 06:46 | there is a cross none as s , there is flux of current movement |
|
| 06:53 | positive current with is within the direction charges within the direction of the |
|
| 07:01 | So as we mentioned in, in first lecture that this ring me and |
|
| 07:06 | of minus 65 millivolts is not something stays locked in as a flat |
|
| 07:13 | it constantly fluctuates and you can have if it tries to depolarize closer to |
|
| 07:21 | we call the threshold for action potential . This value of about minus 45 |
|
| 07:28 | . So this is depolarization in this and in this direction, it would |
|
| 07:33 | hyper polarization and sometimes the membrane may below the what we call the resting |
|
| 07:45 | potential and an abbreviated resting membrane potential RP. So what we have is |
|
| 07:55 | fluctuations. Those fluctuations are dependent on properties, chemical gradient and electrical |
|
| 08:03 | And also some of the thermodynamic changes may be influencing local pieces of the |
|
| 08:07 | . Because when things heat up a bit temperature goes up, ions flocks |
|
| 08:12 | and easier across the membrane too. , the way that this charge is |
|
| 08:18 | is that the negative charge is accumulated the inside of the membrane positive charge |
|
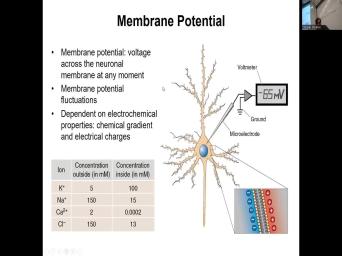
| 08:23 | the outside of the membrane. The of the cell has high concentration of |
|
| 08:32 | . And uh and the outside of cell has high concentration of sodium compared |
|
| 08:38 | the inside uh fluoride compared to the as well as calcium. And as |
|
| 08:45 | can see, the highest difference in , this is the minimal, the |
|
| 08:51 | difference in concentration there exists for calcium their own pile, you have two |
|
| 08:59 | lower on the outside and only 0.0002 the inside, which is 10,000 |
|
| 09:11 | So from pure chemical gradient perspective, would have the highest drive to process |
|
| 09:17 | plasma membrane. But you have to channels. And apart from just having |
|
| 09:22 | that are specific to ions, those must be open and something needs to |
|
| 09:26 | them. And that's something that we're to be learning. So you have |
|
| 09:30 | uneven separation of charge across plasma membrane the inside versus the outside. So |
|
| 09:38 | note that the inner lumen of the and also away from the plasma |
|
| 09:45 | the charges are equal. So the charged area is right around the plasma |
|
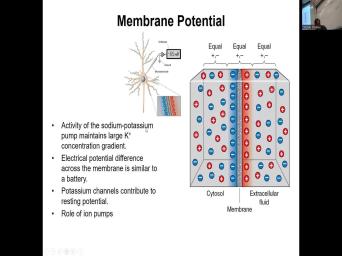
| 09:55 | . Uh It's important again to remind the sodium potassium pumps will maintain large |
|
| 10:02 | concentration gradient on the inside and also con concentration gradients of sodium on the |
|
| 10:10 | . Uh We already talked about electrical . Potassium channels contribute the most arresting |
|
| 10:18 | . And you'll understand why. It's that, that's the way it is |
|
| 10:26 | potentials of ions. So as we spoke that this concentration gradient is not |
|
| 10:33 | controlling how ions are going to cross the membrane through the channels. And |
|
| 10:40 | this example, you have high concentration potassium on the inside of the cell |
|
| 10:46 | you ha have high concentration of sodium the outside of the cell and some |
|
| 10:53 | charged an ion. Then you have channel for potassium here and a channel |
|
| 10:59 | sodium here. And this anion this minus it cannot cross, it doesn't |
|
| 11:06 | a way to cross through plasma So it's trapped, you know, |
|
| 11:11 | the inside or the outside. you can see that sodium starts fluxing |
|
| 11:19 | the area of high concentration and potassium flexing for the area of high concentration |
|
| 11:27 | the area of low concentrations. it never equalizes across plasma numbering. |
|
| 11:35 | you have still more potassium on the and more sodium on the outside. |
|
| 11:42 | that is because as potassium starts crossing the area of high concentration, that |
|
| 11:50 | charge starts accumulating on the outside of membrane, starts accumulating on the outside |
|
| 12:00 | the membrane. And now that positive , the electrical charge starts repelling more |
|
| 12:08 | the potassium ions coming into the area there is less of them. And |
|
| 12:14 | this is called an equilibrium potential. this equilibrium is reached when you have |
|
| 12:25 | channels that have the same amount of that drives concentration and then you have |
|
| 12:33 | same amount, of course, it's and opposite in direction that is driven |
|
| 12:40 | the electrical potential. OK. So is what is referred to as equilibrium |
|
| 12:47 | for each ions. So how do calculate the equilibrium potential? So we |
|
| 13:01 | this nernst equation and nernst equation or potential for ion equation E ion. |
|
| 13:11 | calculated in such a way that each of the ions, each ion has |
|
| 13:20 | own equilibrium potential value. So equilibrium here are calculated for potassium or sodium |
|
| 13:32 | chloride and for calcium are the four ionic species that we're gonna be talking |
|
| 13:39 | . Ek in this case, equilibrium for potassium equals RT over Z fa |
|
| 13:51 | of potassium concentration on the outside of cell versus potassium concentration on the inside |
|
| 13:58 | the cell. R is the gas T is the temperature F is the |
|
| 14:06 | or electrical constant. Z is the charge of an ion. So chloride |
|
| 14:14 | one versus calcium two plus plus And this of course is potassium o |
|
| 14:23 | the outside concentration of potassium on the versus concentration of potassium on the |
|
| 14:33 | OK. So this is again another of this formula and you can read |
|
| 14:39 | on your own. But essentially this 2.303 RT ZF log of ion outside |
|
| 14:52 | ion inside. This whole part of equation collapses into 61 154. And |
|
| 15:03 | value becomes Miller Vault value. And is for monovalent potassium because remember it |
|
| 15:11 | on valence for monovalent potassium, for uh chloride and sodium. But this |
|
| 15:22 | is different for calcium and that's because is divalent to it. So on |
|
| 15:30 | midterm with, with the question commonly I get is am I gonna need |
|
| 15:35 | use a calculator and calculate equilibrium potential to derive the learns equation though you |
|
| 15:43 | not have to do it, but will have to know the concentrations of |
|
| 15:48 | four ionic species on the outside versus inside and be able to easily recognize |
|
| 15:56 | main components and the values equilibrium potential for these four ions that we're talking |
|
| 16:05 | . OK. But essentially, if walk through this calculation here, you |
|
| 16:11 | plug in outside concentration of potassium which low 1 to 20 you'll say what |
|
| 16:19 | this 1 to 20. So if look here the separation of charge is |
|
| 16:23 | there is about 100 millimolar potassium on inside versus five on the outside, |
|
| 16:30 | is 1 to 20. OK. 20 on the inside and one on |
|
| 16:36 | in uh uh on the on the . So this is where this abbreviation |
|
| 16:40 | here. You can use either the millimolar values or the ratio of the |
|
| 16:48 | values for each ion on the outside inside to plug into this uh equation |
|
| 16:55 | . And what you derive is that equilibrium potential for potassium is minus 80 |
|
| 17:03 | . And again, uh just like the rusting membrane potential, some textbooks |
|
| 17:07 | say that resting membrane potential is minus minus 75 minus 65. It kind |
|
| 17:13 | varies depending on the set subtypes of that you're recording in tissues in a |
|
| 17:18 | . But this is uh NS As you can see, you have |
|
| 17:24 | concentration, all ratios of ions outside inside and at 37 °C, which |
|
| 17:33 | your physiological body temperature, you have potential for ions written out here minus |
|
| 17:42 | 62 1 23 minus 65. And show you which ones I I want |
|
| 17:47 | to know because I have a separate that I prepared for you. So |
|
| 17:52 | important points, large changes in membrane potential can result from maybe what |
|
| 17:59 | would call very small and miniscule changes ionic concentrations. So if the changes |
|
| 18:06 | the number and change the voltage fluctuates a bit. That means two things |
|
| 18:12 | that's good because you can produce fast potentials. The other thing is you |
|
| 18:18 | to constantly make sure that the concentrations these ions and substances stay within a |
|
| 18:24 | dynamic range. OK. Net difference electrical charge inside and outside of the |
|
| 18:31 | surface only. So the inside of cell is charged neutral and as you |
|
| 18:35 | further away outside the cell from the , it's also charged neutral rate of |
|
| 18:42 | across membrane. It very much depends the difference between the membrane potential, |
|
| 18:51 | is VM and an equilibrium potential for given ion. And this is what |
|
| 18:58 | call the driving force. This VM is the membrane potential minus equilibrium po |
|
| 19:06 | is what we call the driving Now, concentration difference is known and |
|
| 19:12 | know those concentrations inside and outside of cells, equilibrium potential can be calculated |
|
| 19:19 | you saw here. And so if are different environments where these outside and |
|
| 19:26 | concentrations change, or there are big in temperature somehow, uh these calculations |
|
| 19:34 | equilibrium potential might give you slightly different . So equilibrium potential is not how |
|
| 19:44 | calculate the membrane potential. And that's just now I showed you VM minus |
|
| 19:50 | ion, let's say VM minus And that is because the membrane potential |
|
| 19:57 | on more than just one ion equilibrium calculation. When ernst equation is just |
|
| 20:03 | single ion species. So tacit et Goldman equation is what we use to |
|
| 20:16 | the membrane potential. Now notice this year 6154 millivolts is the same from |
|
| 20:27 | nernst equation. So up to this , we're still 2.303 RT ZF log |
|
| 20:37 | versus inside. Now, let's see that is the same. Now you |
|
| 20:43 | log, you do have the concentration an ion outside potassium. But in |
|
| 20:52 | of that, you have a value is PK and it's not the b |
|
| 21:00 | measure of the substance, but permeability potassium. So again, it's not |
|
| 21:09 | that membranes have channels, those channels to be open. And if they're |
|
| 21:15 | and full, there's going to be permeability for that ion. So there |
|
| 21:20 | to be conditions that create the ability the channels to open and close and |
|
| 21:27 | will regulate the permeability of ions. this is one thing already that is |
|
| 21:33 | from the Nernst equation. The second that is different is that we are |
|
| 21:37 | taking into account more than one And in particular, the most important |
|
| 21:42 | for deriving the DM, which is membrane potential with potassium and sodium note |
|
| 21:54 | at resting membrane potential permeability for this is the PK value for |
|
| 22:03 | It's 40 times that uh sodium, is a resting membrane potential. So |
|
| 22:13 | means that at resting number and potential cell membrane is dominated by potassium |
|
| 22:25 | this is outside, this is inside ions leaking outside the cell. It's |
|
| 22:34 | permeable to potassium press. And I , well, why not sodium? |
|
| 22:39 | because there are certain rules by which channels open and close and also nature |
|
| 22:46 | its own rules and called it potassium channels and they'll be open and therefore |
|
| 22:53 | will be leaking potassium out. And is gonna have the strongest influence on |
|
| 22:59 | resting number and potential because it's gonna 40 times of permeability. So, |
|
| 23:06 | potassium is £40 of permeable, you can plug in their mebolic |
|
| 23:13 | These are the potassium on the outside inside sodium on the outside versus |
|
| 23:23 | And you get the number of potential of about 5 to 65 millivolts. |
|
| 23:34 | now you see the difference between ernst which really is meant to calculate equilibrium |
|
| 23:42 | . It's meant to calculate that electrical chemical force uh interactions and the and |
|
| 23:51 | potential of the membrane. And so values are potential of the membrane |
|
| 24:01 | but they are equilibrium potential values of membrane for each ion. And the |
|
| 24:09 | membrane potential is calculated by taking into permeability for in particular sodium and potassium |
|
| 24:19 | together. OK. To derive this potential in the end. So these |
|
| 24:27 | , as we talked about, they're , they're selective or selectively permeable or |
|
| 24:33 | have selected permeability uh potassium or sodium another ion, but typically they will |
|
| 24:40 | allow other ions to flux through So why is that because they have |
|
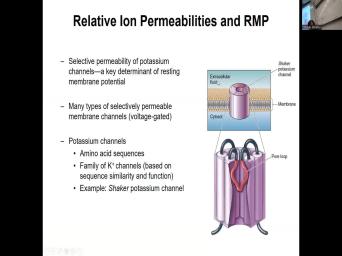
| 24:46 | certain structure and this selective permeability for and having this leak channels is one |
|
| 24:54 | the key determinants of resting member But we have many types of selectively |
|
| 25:01 | membrane channels that are voltage gated that talk about. But we really focus |
|
| 25:06 | sodium of potassium potassium channels. There's whole family of potassium channels that have |
|
| 25:16 | similar sequence of amino acids. Remember are the building blocks from which we |
|
| 25:25 | the channels. And so we can across different channels and we'll see how |
|
| 25:31 | is a significant overlap in the sequences these channels that typically also will select |
|
| 25:38 | potassium. So we will group them the potassium family of channels. And |
|
| 25:43 | will be other sequences that are going be shared among sodium channels and certain |
|
| 25:50 | of allowing sodium current to go And this will be a family of |
|
| 25:55 | voltage gated channels. An example is shaker uh potassium channel. It's a |
|
| 26:04 | interesting channel that if you have a and how it was discovered this potassium |
|
| 26:13 | , a lot of potassium channel structure functionality was discovered by using flies because |
|
| 26:22 | amino acid sequences are not only preserved the same animals but also across different |
|
| 26:34 | , not exactly the same degree. we discover a lot of things and |
|
| 26:40 | primitive organisms like fruit flies and a in the voltage gated potassium channel and |
|
| 26:48 | voltage gated potassium channel was shown to those fruit flies shape. So |
|
| 26:55 | the shaker potassium channel and the shaking the fruit flies was somewhat reminiscent of |
|
| 27:02 | individual having a seizure. And guess a similar type of both educated potassium |
|
| 27:09 | in humans that have an equivalent mutation be found in individuals that have |
|
| 27:19 | And that's how you use a very system, like apply or another like |
|
| 27:26 | side or uh an isolated cell and determine functions and sequences molecular profile. |
|
| 27:36 | then you end up finding the same and equivalent and even symptoms uh in |
|
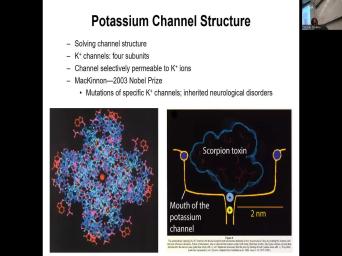
| 27:44 | . So potassium channel structure. So is what Roderick mckinnon, which is |
|
| 27:50 | in your textbook, you have these pathways of discovery and these pathways of |
|
| 27:57 | show you how different individuals used uh knowledge and what they were driven by |
|
| 28:06 | answer big questions in in, in , in this particular uh uh situation |
|
| 28:13 | Neuroscience. And Robert mckinnon, who Nobel Prize in 2003 when mutations of |
|
| 28:24 | potassium channels and study of those potassium and how they're related to inherited neurological |
|
| 28:33 | . Mhm And his story and his of discovery is very, very interesting |
|
| 28:43 | uh I don't remember the details of so you can reread it yourself. |
|
| 28:49 | what I was impressed by is that started out as an as an MD |
|
| 28:56 | had a a really successful practice. then he was longing for basic science |
|
| 29:06 | his medical surroundings. So you decided instead of doing medical practice, he |
|
| 29:16 | going to, he was really driven a question of what is this? |
|
| 29:22 | do these potassium channels look like? was on a quest to see or |
|
| 29:30 | the structure of these potassium channels. it's difficult to do that in humans |
|
| 29:37 | in the clinical setting. So you to enter into basic research setting. |
|
| 29:42 | that's exactly what he did. He out very successful in the career and |
|
| 29:48 | to study fruit flies and mutations and channels and the shaker flies. And |
|
| 29:57 | was using other techniques that are called directed muto Genesis. For example, |
|
| 30:04 | would mutate different parts of this, is depicted as a potassium channel and |
|
| 30:11 | would see which parts of this channel you, when you mutate it change |
|
| 30:16 | function of that channel significantly. For , it can no longer conduct potassium |
|
| 30:21 | it stays open all the time after mutated a certain part of that |
|
| 30:26 | So it's called side directed mutagens. see if I keep right on top |
|
| 30:42 | . So besides directed, OK. to Genesis, that means that you |
|
| 30:56 | directing into a specific site on on this podium channel particularly. So now |
|
| 31:05 | second thing is he was using So nature has a lot of potent |
|
| 31:16 | in it. And this is the toxin, spiders, scorpions, |
|
| 31:26 | reptiles, snakes, they have they have toxins that they produce. |
|
| 31:33 | a lot of these toxins will interact specific channels in our bodies. So |
|
| 31:41 | , you cannot take a scorpion toxin test it on humans. So |
|
| 31:45 | you have to go into a more model like a fruit fly. You |
|
| 31:51 | use these toxins and you can use . Why would you use electrophysiology, |
|
| 32:01 | , electrophysiology will allow you to understand flux of cars across these channels. |
|
| 32:08 | that you're recording electrical activity. So recording membrane potentials, action potentials, |
|
| 32:14 | recording changes in membrane potential. So is electrophysiology then that's cool. So |
|
| 32:32 | exits as an MD and he's using models and he's doing C recognition Genesis |
|
| 32:39 | he's doing electrophysiology and he is using toxins. And all all of these |
|
| 32:46 | are gonna be very helpful for him start deducing the structure and calculating the |
|
| 32:54 | of this voltage gated potassium channel. talking about eighties and nineties, but |
|
| 33:01 | not enough. That's not enough. and he is driven by the quest |
|
| 33:07 | actually visualize the channel. So he leaves this first university that he left |
|
| 33:16 | he was MD and enters into another where he becomes, he becomes an |
|
| 33:33 | first. The longer anybody knows what ray crystallography is to visualize. |
|
| 33:46 | exactly. So it's really complicated. talking about 30 years ago, what |
|
| 33:53 | have to do is you have to and trap one single protean channel inside |
|
| 34:04 | crystal and then you pass x rays that crystal. And those x-rays just |
|
| 34:15 | they show you the bones in the that actually have the ability to show |
|
| 34:20 | the structure. It's called X ray . And to do that, it's |
|
| 34:25 | earning a whole new degree in a different area from side directed neogenesis |
|
| 34:32 | This is in the Department of OK. That's where most of the |
|
| 34:37 | crystallography done. So now he's a . And the reason why uh I |
|
| 34:44 | this is because I think you should these techniques so that you understand that |
|
| 34:49 | that went in in the eighties and nineties and the early two thousands and |
|
| 34:53 | , and not only deriving, calculating structure, understanding important sites in these |
|
| 35:00 | , understanding interaction with the different whether they're blockers, whether they keep |
|
| 35:06 | open, where do they buy? we design now new pharmacological agents that |
|
| 35:14 | these toxins but in a controlled fashion use them as neurological drugs to treat |
|
| 35:22 | disorders, to understand how the structure the function relate together and what agents |
|
| 35:29 | can use to eventually create different medications people that study from uh suffer from |
|
| 35:36 | disorders. OK. So uh the reason why apart from knowing these |
|
| 35:44 | we talked about physiology, talkin will back and talk about it again. |
|
| 35:49 | next lecture or two. So these repeating themes. The other thing that |
|
| 35:55 | really important about this is that if are after something, it doesn't matter |
|
| 36:05 | you are or what degree you have . So as you're embarking on your |
|
| 36:12 | or as you are embarking on your jobs or careers, you have to |
|
| 36:21 | a passion for something. You have want to solve some problem. You |
|
| 36:27 | to improve something, you know, and borer school of business will say |
|
| 36:33 | just wanna make money. But if you make money, it's difficult |
|
| 36:39 | people don't, well, people give money for bad things, but that's |
|
| 36:42 | something that we strive for. So , it's that quest. In this |
|
| 36:48 | , he had a quest of showing structure of the potassium channel MD. |
|
| 36:54 | now phd. This. No, OK. I'll go do the X |
|
| 36:57 | Crystal Large. So it's like reinventing two or three times to answer the |
|
| 37:02 | to finally get the Nobel Prize. his pathway was extremely windy, but |
|
| 37:10 | moving forward. And in this moving forward to, to address a |
|
| 37:14 | , address an issue, solve something innovate. And I think that's, |
|
| 37:18 | really important that you carry that component you and also identify that component within |
|
| 37:27 | field. There are people that are at some things and everybody almost has |
|
| 37:33 | strength that they need to bring out identify and be passionate about it and |
|
| 37:39 | it because otherwise we just can watch on the couch and not much will |
|
| 37:48 | . All right. So it's very that we regulate this potassium. We |
|
| 37:53 | about how the potassium is leaking at . OK. How it's the most |
|
| 38:00 | the membranes of potassium, uh the in number and potential that's closest to |
|
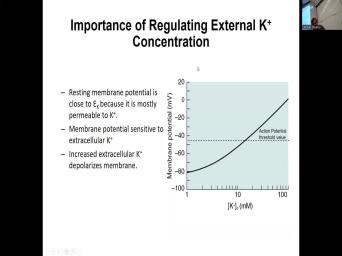
| 38:07 | in C. And if there are fluctuations in the extracellular potassium, this |
|
| 38:13 | influence the overall membrane potential. So measured in millivolts is the membrane potential |
|
| 38:21 | this is ko outside concentration of potassium mila molar. So, a typical |
|
| 38:30 | is about 3.5 5 millimolar which places resting number in potential around minus 70 |
|
| 38:38 | 65 somewhere around here. But if is a change of just five millimolar |
|
| 38:44 | potassium concentration on the outside, this the outside to 10 millimolar, it |
|
| 38:52 | the threshold for action potential generation. means that if that concentration is sustained |
|
| 38:59 | the outside of potassium, the cell fire an action potential. And the |
|
| 39:05 | control of these concentrations of the potassium very important. And if you |
|
| 39:11 | we talked about glia, we talked astrocytes and we said how racy are |
|
| 39:17 | around the neuronal synapses and if there increase in potassium outside, they will |
|
| 39:24 | it up because they don't want the to fire action potentials uncontrollably due to |
|
| 39:30 | changes in the potassium concentration. So it's uh it's, it's |
|
| 39:34 | maintained within a certain range of about millimolar 4.55 millimeter doesn't fluctuate much if |
|
| 39:45 | does, it can actually depolarize the and depolarize them beyond the action potential |
|
| 39:53 | threshold. The other important way that regulate potassium. So this is the |
|
| 40:01 | , the next slide of how they sensitive increases in exercise of the potassium |
|
| 40:06 | they will essentially fally buffer this potassium their processes. And the interconnective network |
|
| 40:13 | other troys also from the blood. , we have regulation of both endothelial |
|
| 40:19 | t junctions and Astros on how much from the blood can enter into the |
|
| 40:26 | . And meaning that if there is much of potassium entering into the brain |
|
| 40:31 | the external environment, that means the is increasing, the brain can become |
|
| 40:36 | excitable and that could be due to leaky blood brain barrier and high concentrations |
|
| 40:41 | potassium in the blood. All So this concludes our lecture on the |
|
| 40:48 | number and |
|