| 00:00 | Welcome. Uh this in this we're going to continue on with metabolism |
|
| 00:06 | build on what we learned previously uh respiration. So, recall respiration uh |
|
| 00:12 | to that um process is electron transfer recall the functioning of electron transfer uh |
|
| 00:22 | uh embedded in the membrane. Uh presence of a donor to as a |
|
| 00:26 | of electrons that then flows through the transport chain, using that energy to |
|
| 00:32 | pump protons out generate a proton Uh electrons flow into a term acceptor |
|
| 00:38 | then um the the energy from the being used to produce a TP. |
|
| 00:43 | in this, we're gonna really look a little more detail at that |
|
| 00:48 | And so in recent years, we've been uh discovered that uh that electron |
|
| 00:57 | , although we're familiar with the process transfer occurring within a single cell from |
|
| 01:01 | donor to an acceptor, that this also occur among uh species within species |
|
| 01:08 | from one species to another species. so a geo backer is was the |
|
| 01:14 | is the uh strain that's been most in this uh aspect in this |
|
| 01:19 | So it's called interspecies electron transfer. not within a single cell but from |
|
| 01:25 | a cell of one species to that another. Right. So you see |
|
| 01:29 | in both cases, what's um of course, electron transfer process is |
|
| 01:37 | presence of a of a donor, , electron donor. In both cases |
|
| 01:48 | and here acetates serves as a donor then uh electrons are transferred to another |
|
| 01:56 | . And so we have a terminal at the end, in this |
|
| 02:07 | nitrate reduced to ammonia co2 to uh . And so this is features of |
|
| 02:19 | electron transfer system. Uh And, the unique here is the fact that |
|
| 02:24 | are um transfer electrons through different OK. And uh in essence completing |
|
| 02:32 | electrical circuit, if you will, now know that. So here we |
|
| 02:36 | a conduct a conductive material um between two species that serves to facilitate electron |
|
| 02:43 | . Of course, also, it's discovered that these bacteria have these |
|
| 02:48 | right, specialize for electron transfer like wires in essence. So, uh |
|
| 02:55 | , and further, we found that , these can also form biofilms. |
|
| 02:58 | it can be what's called an electrogenic that conducts electricity. So very, |
|
| 03:03 | unique in finding. Um there are are various applications for these uh |
|
| 03:08 | in different ways. So, but point here is is uh we're gonna |
|
| 03:13 | on this process of electron transfer and components involved. OK. So that's |
|
| 03:19 | of course, meaning we're gonna talk um redox reactions, right? So |
|
| 03:25 | uh in the process of electron of course, uh become uh |
|
| 03:30 | become alternately become oxidized and reduced uh they uh receive and then uh give |
|
| 03:37 | electrons uh looking at the component electron chain. Um uh And then the |
|
| 03:43 | of reduction potentials, what will begin , right, because molecules have different |
|
| 03:49 | infinities in terms of how well they uh receive electrons and give up |
|
| 03:53 | So there's differences in those abilities, molecules are better at donating electrons, |
|
| 03:58 | are better at accepting electrons. Um then uh we'll look at it a |
|
| 04:02 | bit at uh comparing aerobic versus aerobic as well. OK. So, |
|
| 04:09 | so again, we're, this we're talking about here is respiration uh |
|
| 04:13 | relies on the electron transport system. . So, um uh so organic |
|
| 04:18 | is that um is that uh process gonna look at? OK. Uh |
|
| 04:26 | also as well. Um reservation can um with a organic substrate as the |
|
| 04:33 | donor or inorganic substrates as the That's the difference between uh organic troph |
|
| 04:39 | you will and the little little it's an inorganic source. So we'll |
|
| 04:44 | more at lits in the part two chapter four, right? So, |
|
| 04:48 | this first part, we're gonna look uh really or or organic tropes. |
|
| 04:51 | , using those that use organic Uh uh um so, let's just |
|
| 05:00 | real quick, just kind of an . And we saw this uh we |
|
| 05:03 | at this last time is the in uh in um respiration utilizing electron |
|
| 05:11 | system, the membrane is essential. the membrane contains the uh electron transport |
|
| 05:17 | . Um There's a source of electrons feed the electron transfer chain. It |
|
| 05:22 | be organic or inorganic. OK. the difference between organic tropes and |
|
| 05:27 | That source is going to be electron . So hence, it's reduced and |
|
| 05:32 | become oxidized, giving up electrons. I remember the uh electron carriers that |
|
| 05:37 | universal in such systems. So N uh also, of course, we |
|
| 05:42 | F AD as well. So these carriers then become reduced in the process |
|
| 05:49 | they're the ones that interact with electron chain giving up electrons. And then |
|
| 05:53 | alternately, you know, passing them through the chain. And so to |
|
| 05:59 | terminal acceptor that becomes reduced. And we, we're maintaining electron flow as |
|
| 06:04 | do this. OK. So, there can be uh it's aerobic |
|
| 06:08 | oxygen, of course, something other oxygen. It's anaerobic respiration and that |
|
| 06:14 | will become reduced in the process as receives electrons, right? So it's |
|
| 06:19 | about redox reactions. OK. Oxidation . And so the the flow that |
|
| 06:27 | learn, we'll learn that the flow maintained um by setting the system up |
|
| 06:33 | that we begin with very strong electron and then sequentially getting progressively more and |
|
| 06:40 | stronger acceptor. And that's what enables flow to occur, right. So |
|
| 06:47 | , so as you might guess the module here at the beginning, it's |
|
| 06:55 | be a very strong donor, strong . OK. Strong electron donor. |
|
| 07:08 | . And this of course is gonna a very strong accept her. And |
|
| 07:18 | oxygen has what's called the highest reduction . OK. It's a very good |
|
| 07:25 | , at receiving electrons. Uh And setting up setting up the system this |
|
| 07:30 | , strong donors to progressively stronger and acceptor electron flow is maintained. And |
|
| 07:35 | of course, is used to energy that is used to pump protons out |
|
| 07:39 | generating a proton motive force and then the energy um as protons flow that |
|
| 07:47 | to produce uh lead to the formation A TPS. So this is |
|
| 07:53 | you know oxy, the phosphorylation is chem osmosis mechanism we'll talk about |
|
| 07:58 | OK. So really in a that's really what we're gonna be focusing |
|
| 08:01 | is are different aspects of this OK. And so the overriding concept |
|
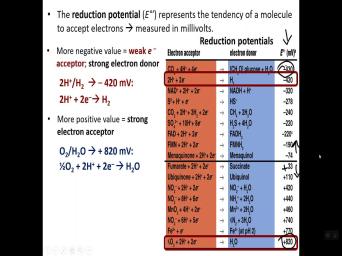
| 08:08 | is reduction potential, right, redox potential, right. So uh the |
|
| 08:14 | is reduction potential and of course, equates to delta G as well. |
|
| 08:19 | we'll see how those two correlate. . So when we look at the |
|
| 08:25 | potential, there is, there's a that's given for these, that's called |
|
| 08:30 | electron tower is typically what they're And so we, we look at |
|
| 08:35 | , the a ranking if you will different reactions, molecules involving different reactions |
|
| 08:42 | their ability as an electronic sector. you see the red block there, |
|
| 08:46 | looking at reactions in which it's it's a molecule receiving electrons and becoming uh |
|
| 08:53 | . OK. And so of each of these has an electron accept |
|
| 08:58 | and an electron donor reform, And then of course, there's a |
|
| 09:01 | reduction potential value on the far right . OK. Here that, that |
|
| 09:10 | a uh uh a certain magnitude, it's a negative per uh at the |
|
| 09:15 | of the uh table, uh very values to values at the bottom of |
|
| 09:21 | table which are very positive. So we're going from negative to positive |
|
| 09:27 | as we go from top to OK. And so this equates to |
|
| 09:32 | different things and I have drawn a in the middle that separates um kind |
|
| 09:38 | a dividing line between negative values and values. OK. And so, |
|
| 09:47 | if you look at an example of couple of these reactions, right? |
|
| 09:51 | the thing to remember is the values are more negative are those that we |
|
| 09:56 | to be strong electron donors, So if they're a strong donor, |
|
| 10:01 | gonna mean conversely that they're a weak . OK. Conversely, those that |
|
| 10:08 | more positive values such as oxygen are strong electron acceptor. Um And |
|
| 10:15 | uh the, the, again, potential is, is that tendency of |
|
| 10:19 | of a molecule to accept electrons. remember if you're accepting electrons, you're |
|
| 10:23 | reduced, hence the term reduction And so those that are very positive |
|
| 10:29 | are very strong acceptor. OK. those are the kind of molecules you're |
|
| 10:34 | see that serve as the terminal electron in the respiratory chains. So oxygen |
|
| 10:42 | iron, uh iron three plus uh nitrate. These are common to be |
|
| 10:50 | the end uh in in whether it's respiration or anaerobic respiration at the terminal |
|
| 10:55 | . OK. Strong electronic acceptor And so here's an example of a |
|
| 11:00 | donor. OK. Or a weak , right? So you're looking at |
|
| 11:05 | proton hydrogen couple. OK. And uh the the the reaction of protons |
|
| 11:14 | electrons OK? To produce hydrogen that's a very negative reduction potential minus |
|
| 11:22 | 20 millivolts. OK. So that protons are not very good at accepting |
|
| 11:29 | , right? Conversely, the oxygen couple, right? Plus the |
|
| 11:35 | the most positive value reducing potential plus 20 millivolts. And so uh that |
|
| 11:43 | is uh oxygen and protons. Uh receives electrons to, to form |
|
| 11:49 | right? Very positive. So what that mean in terms of negative and |
|
| 11:53 | values or reduction potential that equates to delta G value? OK. So |
|
| 11:59 | delta G uh processes that are negative G release energy, right? Those |
|
| 12:06 | are positive delta G require energy to proceed. OK. And so |
|
| 12:13 | native delta G values equate to positive potentials. OK. So you can |
|
| 12:21 | there what this will tell you is water. I'm sorry, using |
|
| 12:26 | using oxygen as a terminal acceptor can to a very negative L to G |
|
| 12:38 | , right? Using protons as electron would be a positive delta G |
|
| 12:50 | Hence that that's why we look at as not being favorable in the role |
|
| 12:54 | electronic sector. It's very weak because actually would require an input of energy |
|
| 13:01 | , to, to do that. . Whereas um using oxygen as an |
|
| 13:09 | question, more uh favorable because it uh result in the release of |
|
| 13:15 | right? So that's how we equate two. What we label it as |
|
| 13:19 | , a weak acceptor or a strong . OK? And also remember that |
|
| 13:25 | something is a weak acceptor, it conversely then be a strong donor and |
|
| 13:29 | versa. OK. So let's look a couple of examples here. Uh |
|
| 13:35 | we'll stick with the same uh So redox couples, right? So |
|
| 13:38 | gonna combine. So the thing I remember of course is something's something's being |
|
| 13:43 | and it's gonna then something has to oxidized at the same time, |
|
| 13:46 | So it's always gonna be AAA donor an acceptor. So redox always occurs |
|
| 13:52 | . And so what, what, we try to do in electron transfer |
|
| 13:55 | and setting of electron transfer system is arrange donors and acceptor such that it |
|
| 14:03 | give us a favorable delta G, ? A a net negative delta G |
|
| 14:07 | that will release energy. So you an electron transfer system that is, |
|
| 14:13 | is optimal is one that is releasing , right? Because we're gonna use |
|
| 14:16 | energy to pump protons out. So it all relates back to this |
|
| 14:22 | mode of force we're gonna be talking here shortly. OK. So here |
|
| 14:28 | again our example of our uh protons electron acceptor, which is a very |
|
| 14:33 | ex acceptor. Um But we can look at look at it from the |
|
| 14:38 | of the donor, right? So gas as a donor, right? |
|
| 14:43 | when we do that, we look the reverse reaction, we will change |
|
| 14:47 | magnitude of the reduction potential, So now looking at hydrogen gas as |
|
| 14:55 | donor, that is a positive reduction . OK. So um so using |
|
| 15:06 | as a electron donor is very right? Because you know that positive |
|
| 15:11 | potential equates to a negative delta OK. So if we set up |
|
| 15:15 | electron transfer system where we have hydrogen a donor, strong donor and then |
|
| 15:22 | as the acceptor, we already know a strong acceptor. So now |
|
| 15:28 | we're, we're, we're a system we have a strong donor at the |
|
| 15:31 | and a strong acceptor at the OK. And now we have a |
|
| 15:36 | uh reduction potential that will give us very favorable negative delta G. |
|
| 15:42 | So remember that, that energy release going to, is going to work |
|
| 15:47 | , right? So we're gonna have flow, right? And we'll be |
|
| 15:50 | to pump protons out and then get energy back in the form of a |
|
| 15:54 | . So that's, that's the, the why that, that's why these |
|
| 16:01 | are set up in this fashion, donor to strong acceptor because that's a |
|
| 16:05 | , gives us a very favorable um potential, positive reduction potential, |
|
| 16:11 | which equates to a uh a very negative delta G and energy release that |
|
| 16:16 | can do something with. OK. again, the other thing you remember |
|
| 16:20 | the reversibility, right? So if look at a, a system where |
|
| 16:23 | appears to have a very well does a sort of a very weak electron |
|
| 16:28 | ie protons. But if you look the uh at the, at the |
|
| 16:33 | form of that reaction, the hydrogen and how is it, you |
|
| 16:37 | how would it be as a, a donor? We flipped a |
|
| 16:40 | reverse it and then we flipped the of the reduction potential. So it |
|
| 16:44 | out that, you know, protons very poor as being an acceptor, |
|
| 16:48 | hydrogen is excellent at being an electron in terms of energetics. OK. |
|
| 16:53 | that's what it's all about. So , uh let's look at another |
|
| 17:00 | of course, in, in uh respiration uh N A DH right before |
|
| 17:05 | A DH in the course of glycolysis through in, in the um uh |
|
| 17:12 | oxidation uh to and then to the cycle. So, remember we're |
|
| 17:17 | we accumulate lots of N A right? And so that's what serves |
|
| 17:22 | the uh donor to the, the direct donor to electron transfer |
|
| 17:26 | So we look at the reaction of , right, uh to, to |
|
| 17:33 | reduced to ne DH that actually is a, it actually is a poor |
|
| 17:39 | acceptor. Um But the N A form, that is the electron carrying |
|
| 17:47 | that uh becomes oxidized. So that's very good donor, right? So |
|
| 17:51 | , we, we do, we the reaction and we change the magnitude |
|
| 17:55 | our sign. And so um we uh then of course, now it's |
|
| 18:02 | plus 3 20 millivolt deduction potential which to a negative delta G. |
|
| 18:06 | yes, al although Ned the formation N A DH from the reduction of |
|
| 18:11 | is energetically not favorable, remember that ad acts as a co factor with |
|
| 18:16 | . And so that combination can, , of course, uh um um |
|
| 18:23 | the, the not so good And, and of course, the |
|
| 18:27 | go works because we go of lots of an A DH. But |
|
| 18:32 | an A DH as electron donor to system is very good because we're gonna |
|
| 18:35 | that now with oxygen as a term . OK. And so when we |
|
| 18:41 | at that and then combine them, all additive. And so we get |
|
| 18:45 | net uh very good net uh energy a very good negative delta G using |
|
| 18:52 | A DH as a donor auction as acceptor. And of course, the |
|
| 18:57 | is used to put protons out, a proton gradient. Then we're gonna |
|
| 19:01 | that energy back through the formation of TP uh through the uh che osmosis |
|
| 19:07 | as we'll talk about shortly. So, um so of course, |
|
| 19:13 | we look at uh bacteria nature and know what are the and those that |
|
| 19:17 | of course respire whether aerobically or they of course, uh combine can |
|
| 19:25 | different electronic donors and electronic acceptor. course, it depends on their genetic |
|
| 19:30 | and what's available to them and the in terms of nutrients. And so |
|
| 19:34 | can, we can use, you , the data from the table to |
|
| 19:38 | of speculate. Uh would it be favorable to use electron donor? X |
|
| 19:44 | , and combine it with electronic Y, would it work? And |
|
| 19:48 | is what this question is asking, a bacterium obtain energy from Xin as |
|
| 19:53 | electron donor and nitrate as an electronic ? It's easy enough to, to |
|
| 19:59 | out using the values from the And what we know about how we |
|
| 20:04 | at electron acceptor electron donors. So here's the, the part of |
|
| 20:09 | table that corresponds to this. So as a donor and nitrate as an |
|
| 20:14 | . So remember that the tables are , you know, the left column |
|
| 20:18 | all as electronic acceptor, right? we're gonna have to do some |
|
| 20:22 | So here's so the first thing step to set it up as we're looking |
|
| 20:25 | this is the question that we're asking as a donor suckin becomes oxy |
|
| 20:31 | And those electrons are then given up electron transport chain. They're looking at |
|
| 20:37 | as an acceptor, nitrate reduced to , right? So that's our, |
|
| 20:41 | our system. So then we have look at su as the donor, |
|
| 20:46 | right, oxidized to fate. And that's going to change the magnitude of |
|
| 20:53 | reduction potential from a plus to a , right? And that actually equates |
|
| 20:57 | a pos positive delta G and we oh That may not work so |
|
| 21:00 | OK. But it depends on how nitrate is right as an except as |
|
| 21:05 | donor, I'm sorry, as a as an acceptor. And so here |
|
| 21:10 | have that reaction. So it doesn't . So it's it's gonna stay as |
|
| 21:14 | in the table nitrate reduced to uh trite. And uh that's a very |
|
| 21:22 | reduction potential, very positive reduction which equates to a very uh a |
|
| 21:27 | delta G. OK. And so look at the net, so these |
|
| 21:34 | additives. So we look at the result, net reduction potential is a |
|
| 21:37 | . That's still gonna equate to a good negative delta G. So |
|
| 21:41 | that can work even though is not in terms of reduction potential, having |
|
| 21:48 | slightly positive uh um uh uh positive G, it, it will work |
|
| 21:56 | we're combining with a very, very strong uh acceptor nitrate. |
|
| 22:01 | And so, um and so that can any combination of donor acceptor receive |
|
| 22:06 | en favorable in this way. So, uh so the key here |
|
| 22:11 | really just to look at. So asking suck as a donor. So |
|
| 22:15 | on this side. So we just to flip it around right here. |
|
| 22:23 | as we do, we're going to that. OK. Um But |
|
| 22:29 | whatever we're combining it with uh it's additive. And if the net |
|
| 22:33 | is deposit reduction potential that equates to negative delta G, OK. |
|
| 22:38 | and it's favorable, right? All . So if we uh look at |
|
| 22:45 | components of a electron transport chain, . So, um again, they're |
|
| 22:53 | be in the membrane, first and , they're gonna be stuffed into a |
|
| 22:56 | for bacterial cells. This will be uh in the plants in the, |
|
| 23:00 | the cell, cyto plastic membrane. . And again, we're gonna arrange |
|
| 23:05 | components from strong donor to strong And of course, the typical components |
|
| 23:10 | the electron transfer chain are cytochrome, very large molecules containing hem groups, |
|
| 23:16 | you see there, uh uh uh sort of a metal, typically iron |
|
| 23:22 | the um uh atom in the middle the molecule or in the middle of |
|
| 23:26 | , of the ring ring to, ultimately accept and, and give up |
|
| 23:32 | . Uh could also be things like atoms or common copper atoms. Uh |
|
| 23:36 | are the, the, the centers , where the reduction oxidation reductions |
|
| 23:41 | OK. Hence, we call these reduct cases because they, they alternately |
|
| 23:46 | and then give up electrons. There's typically those shuttle or small organic |
|
| 23:51 | I think electron shuttles between these larger that we call um uh what we |
|
| 23:58 | quins um as we will see. so, um so electron flow, |
|
| 24:04 | ? So we're going from strong donor strong separate reduction potential is increasing, |
|
| 24:09 | ? We're going from a more negative more positive, right? And that's |
|
| 24:16 | enables electronic flow to go from donor acceptor, right? And of |
|
| 24:20 | this is a company that's gonna be negative delta G and that energy can |
|
| 24:25 | used to pump protons out. So, and of course, don't |
|
| 24:34 | forget that we're looking at aerobic respiration , it could easily be nitrate as |
|
| 24:39 | uh a terminal acceptor for me, or some other uh nonoxynol if it |
|
| 24:46 | anaerobic respiration. OK. So, here we see uh the, the |
|
| 24:53 | system in E coli uh this will under a aerobic conditions from E |
|
| 24:58 | It can do a number of, can also respire anaerobically. So it |
|
| 25:01 | depends on what's available to it. If it were anaerobic conditions, it |
|
| 25:05 | express the particular genes for the particular cytochrome that would interact with nitrates if |
|
| 25:10 | were doing nitrate respiration. But so we're gonna have um um an |
|
| 25:17 | uh what's called oxo or duct Here. It's uh an A |
|
| 25:22 | the hydrogenate that will interact with N DH from a, from a number |
|
| 25:27 | N A DH molecules in the process glycolysis and soul aspiration as you see |
|
| 25:35 | . Um And then these will interact uh react with N A DH D |
|
| 25:40 | and that's where the uh oxidation that D occurs. Electrons are and received |
|
| 25:46 | that complex. Uh This is a point where protons are formed or |
|
| 25:51 | up, excuse me. And then are the electron shuttles, right. |
|
| 25:56 | quinones become um are initially oxidants become , receiving electrons from the N DH |
|
| 26:03 | dehydrogenase and then pass those on to . Uh And there can be more |
|
| 26:08 | one cytochrome. And so uh there's terminal cytochrome oxidase that will interact in |
|
| 26:13 | case with oxygen. So we're looking aerobic respiration, here's oxygen here. |
|
| 26:19 | um again, the pumping of all right. So these are |
|
| 26:24 | these electron transfers uh are coupled to pumping of protons at these two |
|
| 26:32 | OK. So we're generating that proton as a result of these electron |
|
| 26:36 | OK. And so of course, , the, the levels of these |
|
| 26:40 | can change depending on the, you , the environmental conditions. Is it |
|
| 26:44 | it, you know, is it oxygen? Is it higher oxygen |
|
| 26:47 | Is it no oxygen? And then can change the cyco completely in order |
|
| 26:52 | be able to interact with the proper terminal acceptor. So uh so these |
|
| 26:56 | can change depending on uh environmental So, uh but the point is |
|
| 27:03 | having elect Trump flow, of going in this fashion, OK, |
|
| 27:10 | a terminal acceptor donor, to accept strong donor to a strong terminal |
|
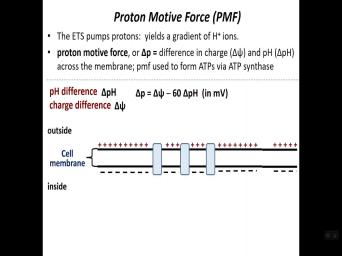
| 27:15 | Um And so the pro time out force, right? So this is |
|
| 27:19 | chemi osmosis mechanism. So there's 22 involved. OK? There's um a |
|
| 27:28 | because we're dealing with protons, And we are forming a gradient of |
|
| 27:34 | because one of the membrane essential in right electron transfer components are in the |
|
| 27:38 | and the energy from the transfer electrons used to pump protons out. So |
|
| 27:44 | creating a gradient. So hence, , we're, we're, we're concentrating |
|
| 27:47 | on the one side of the which means there's gonna be a P |
|
| 27:51 | because P equates to hygenic concentration, ? There's gonna be a Ph |
|
| 27:57 | But because protons are charged right, also gonna be a charge difference. |
|
| 28:01 | that's so delta ph and then delta is a charge difference. OK? |
|
| 28:06 | so the, the almost all most cells uh are are negative with respect |
|
| 28:16 | the intracellular side of the membrane versus exterior side of the membrane. And |
|
| 28:20 | really due to the to the presence proteins in the cell. Uh proteins |
|
| 28:24 | very large molecules. So they don't easily, they're not gonna pass, |
|
| 28:30 | know, II I and, and through membranes, obviously. So the |
|
| 28:35 | inside the cell are pretty, pretty are staying there for the most part |
|
| 28:38 | they are a type of protein to secreted. But there's gonna be lots |
|
| 28:42 | proteins in the cell and that contributes really to the negative charge that's inside |
|
| 28:45 | a cell. And so because the grating generates positive charge outside the |
|
| 28:50 | there's gonna be this um this difference charge across the membrane, positive outside |
|
| 28:56 | inside. OK. Um yes, are other types, many other types |
|
| 29:00 | ions that contribute to charge. But overwhelmingly the internal charge is really the |
|
| 29:05 | that are the biggest contributor there. uh so our equation then we got |
|
| 29:10 | pro time amount of force. So P is pro time amount of force |
|
| 29:14 | that difference in charge, delta And the difference in Ph. And |
|
| 29:19 | the equation there is uh uh delta sign minus 60 times delta ph. |
|
| 29:24 | . And so uh the protons then electron transfer energy for electron transfers generates |
|
| 29:35 | the page difference, the charge So hypothetically we've put in here uh |
|
| 29:40 | difference of 6.5 outside P to 7.5 . So it's delta ph of, |
|
| 29:46 | one. OK. And so then have the A TP. So the |
|
| 29:53 | to remember is that, and the motive force comes from this concentration |
|
| 29:58 | OK. And the attraction of those charged protons to the negative charge inside |
|
| 30:07 | cell. So we've, we've created difference of concentration difference of protons outside |
|
| 30:16 | inside, which means they would gladly back into the cell if they're given |
|
| 30:21 | way to do. So two, also have the charge traction positive to |
|
| 30:27 | charge. So we have that So there's two forces the force for |
|
| 30:32 | to move, slow down their gradient the cell plus the charge traction, |
|
| 30:37 | them into the cell. And but being a charged ion, they can't |
|
| 30:43 | truly diffuse through yourself. They only very, very slowly if at |
|
| 30:48 | And so if we provide them a way, then we can then harness |
|
| 30:53 | proton mode of force. And that's the A TPS, that's the role |
|
| 30:58 | those. So our, our A is for short, this is how |
|
| 31:01 | going to um use the energy from , the flow of protons down their |
|
| 31:08 | . OK. So they'll flow through the gradient as well as being attracted |
|
| 31:13 | that negative charge. And that's what the proton mode of force. The |
|
| 31:17 | is used to form a TPS. . So if we just look at |
|
| 31:22 | typical uh range of charge that's in typical ee coli cell or any bacterial |
|
| 31:29 | for the most part, it's in range of minus 50 to minus 1 |
|
| 31:33 | millivolts. So, if we have delta ph of one, the range |
|
| 31:36 | values is anywhere from a motive force minus 1 10 millivolts to minus |
|
| 31:43 | 10 millivolts. That's kind of the average range um of, |
|
| 31:48 | of the proton motor force. And of course, remember, the |
|
| 31:52 | motive force can be used for things than strictly for uh a TP |
|
| 31:58 | They can also be directly tied to a flagellum, for example, uh |
|
| 32:02 | move uh other molecules in through different transport proteins. So the the proton |
|
| 32:08 | force here is used for uh cellular other than uh A TP formation. |
|
| 32:15 | obviously, that's an important, important function, but there are other functions |
|
| 32:19 | well for this, for this And the, and the eco does |
|
| 32:22 | it for those purposes as well as other bacteria obviously. So, um |
|
| 32:29 | if uh so again, the, key things about this proton force is |
|
| 32:33 | it's in a membrane, it relies electron transfer uh chain embedded in |
|
| 32:38 | The membrane is what creates two sides uh to which you can concentrate protons |
|
| 32:44 | one side. Um Then there's a difference and so protons are char are |
|
| 32:49 | also to the negative charge inside the as well as the concentration difference as |
|
| 32:54 | go down their gradient, they release , right? So remember that a |
|
| 32:57 | concentration gradient is a is a form potential energy. And this pro and |
|
| 33:03 | any molecule moves down their gradient higher lower they release energy, right? |
|
| 33:09 | uh and that energy here is in case, we see in this example |
|
| 33:12 | to form a TPS. OK. um and and again, just to |
|
| 33:20 | to, to make the whole thing of course, electron transfer chain, |
|
| 33:22 | electron donor need internal acceptor. So all this kind of goes hand |
|
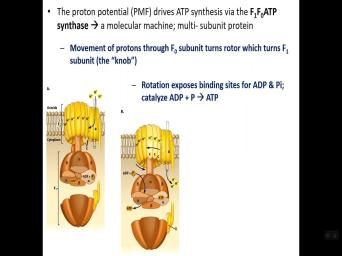
| 33:27 | hand. All right. So looking a little closer at the A TP |
|
| 33:35 | , OK. Um The, this a very large protein multi protein |
|
| 33:44 | OK. It's, it's an example a of a nano motor if you |
|
| 33:49 | because it does, it does rotate it's all through of course, the |
|
| 33:56 | presence of a proton gradient that makes run because protons will flow through the |
|
| 34:03 | and that flow generates a movement of complex. OK. So we have |
|
| 34:10 | parts what's called an F sub zero F sub one. So the protons |
|
| 34:17 | through components uh the component in a , in the F subzero which is |
|
| 34:21 | in the uh cystic membrane. The portion if you will or the mob |
|
| 34:28 | call it is is below the membrane the cytoplasm. OK. There are |
|
| 34:36 | sites within the F uh sub one . As you see here, there's |
|
| 34:44 | 80 binding site for A P and . OK. And we're gonna look |
|
| 34:53 | the animation and kind of illustrates this . But as ad P and phosphate |
|
| 34:56 | , as the rotor turns, the it ultimately exposes sites in the F |
|
| 35:07 | subunit. OK. And when those up, that's when A TP and |
|
| 35:10 | can bind and then as they that provides energy to form a |
|
| 35:16 | OK. Then the A TP will eventually release as the rotor moves and |
|
| 35:23 | an opening. Such the A TP unbind and release. So there's ad |
|
| 35:27 | and phosphate going into the complex att and it is then released and then |
|
| 35:32 | be used to do or whatever, the need is for A TPS. |
|
| 35:37 | . But it's a complex that's moving , and, and generating this binding |
|
| 35:42 | unbinding. Uh There can be, course, uh sodium pumps that, |
|
| 35:50 | don't rely on proton gradient, but on sodium gradient. Uh of |
|
| 35:54 | halo phys that live in very high conditions uses. But also there are |
|
| 35:58 | pathogens because uh sodium ions are very in the human body. And so |
|
| 36:04 | have evolved ways to sodium pumps in same manner to, to generate a |
|
| 36:09 | uh formation. So let's look at animation here. OK. So here's |
|
| 36:18 | , our um A pas F zero one. There's our protons that have |
|
| 36:25 | generated proton gradient. And so as um sorry, so here's a sliced |
|
| 36:36 | cutaway view of the F one sub and in the F zero, so |
|
| 36:44 | the F zero recipient. So as flow through it turns and then |
|
| 36:51 | it then turns the rotor and what's called the knob at the |
|
| 36:55 | the F one uh unit. And can see how a TPSATP and phosphate |
|
| 37:02 | in a TP is formed and released the rotor is turning. OK. |
|
| 37:08 | comes ad P and phosphate, ad released. So let's take a look |
|
| 37:12 | , look at a cross section through complex. So we can see that |
|
| 37:20 | rotor is not a complete circular There's actually it has like a teardrop |
|
| 37:26 | with a little pointed end and you the alpha beta units of the F |
|
| 37:31 | complex. And so here we'll see , you see AD P and phosphate |
|
| 37:37 | here here it's bound. Uh And a TP is gonna be formed as |
|
| 37:42 | result and then it will be And so if you look at the |
|
| 37:47 | of the axle is called, all . So you can see in that |
|
| 37:51 | where the pointed end is exposing so the A TP can unbind and |
|
| 37:57 | then as the rotor moves. All . So now the space is open |
|
| 38:01 | AD P and phosphate to enter and as the axle rotates like so, |
|
| 38:09 | you can see at the upper top , that energy is used to form |
|
| 38:14 | TPS. All right. So this gets released and then the other one |
|
| 38:19 | in road or moves that a TP become released. But as it |
|
| 38:24 | then the energy from that is then to form a TPS here. So |
|
| 38:29 | alternate. So as a rotor we alternately um unbind, formed a |
|
| 38:34 | , bring in a TP and phosphate form a TP. So this all |
|
| 38:40 | alternately as this rotor is turning to generate of course, lots of |
|
| 38:45 | TPS uh as protons flow through. . So, um and so |
|
| 38:52 | what keeps it going, of course having electron transfer chain having the donor |
|
| 38:58 | becoming. Um uh so we have sort of electron source, right? |
|
| 39:03 | have the glucose as a source, ? We oxidized glucose form any ed |
|
| 39:07 | ned gets reduced to nd those N S go to electron transfer chain uh |
|
| 39:13 | oxidized and electrons flow through the electron chain to a internal strong to a |
|
| 39:18 | acceptor, strong donor to strong Uh that gives a nice delta G |
|
| 39:23 | energy is used and the pump protons protons flow to a TP A to |
|
| 39:28 | generate a TP. So it all together, right? All of course |
|
| 39:32 | with electron donor feeding electron transfer chain a term acceptor and the pump |
|
| 39:39 | So it's all, it's all right. Uh So if we then |
|
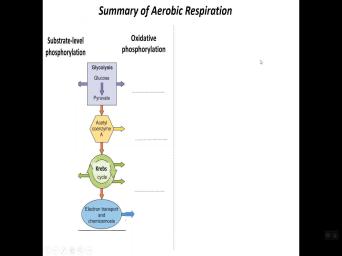
| 39:46 | at kind of a summary of a , right. So here the stages |
|
| 39:51 | talked about previously glycolysis uh formation crypt . So of course, remember the |
|
| 40:00 | level phosphorylation, we have a couple steps in glycolysis in a crypt cycle |
|
| 40:04 | that happens. But through ox we form lots of N A DH |
|
| 40:09 | at each stage. And then of , fa DH and crypt cycle as |
|
| 40:14 | . So we form, so we we have a number of these uh |
|
| 40:17 | electronic carriers in ad fa DH And it's been the, the, |
|
| 40:23 | , you know, determinations have shown for about uh eight protons are pumped |
|
| 40:28 | each and a DH oxidized. So get one A TP for about three |
|
| 40:34 | . OK. And again, these average values and so this equates to |
|
| 40:41 | almost three A TPS per N A oxidized and about 1.5 a TPS for |
|
| 40:47 | fa DH two oxidized. And the there is simply because uh the oxidation |
|
| 40:51 | N A DH is linked to two pumping mechanisms. Fa DH is only |
|
| 40:57 | to one. So there's gonna be little bit difference in energy output. |
|
| 41:02 | And so if we do the then we produce 10 A DNA DH |
|
| 41:06 | that gives us approximately 27 A TPS for two FA DH S two is |
|
| 41:11 | , it gives us three approximately. a total of 30 A TPS via |
|
| 41:18 | via oxidative phosphorylation, right. This all oxidative phosphorylation and then occurs. |
|
| 41:25 | 30 versus four right through substrate level . So there's a big difference, |
|
| 41:33 | . And in reality, I that's a theoretical, you know, |
|
| 41:37 | approximately. In reality, the levels actually a little bit lower than |
|
| 41:42 | It can be like in the low mid twenties, uh maybe 18 or |
|
| 41:46 | because the the proton motor force is for things other than making a |
|
| 41:51 | And so there's gonna be different different demands on sell at different times |
|
| 41:55 | maybe producing more TPS. Uh The amount of may may fluctuate that are |
|
| 42:00 | formed using a proton amount of Uh because it's used for transport of |
|
| 42:05 | molecules to move and things like So that's kind of why it, |
|
| 42:09 | vary. Um But you know, obviously it's a much higher level a |
|
| 42:15 | than it is through substrate level So we look at anaerobic respiration. |
|
| 42:22 | . Um There can be um um course, a number of animal reservation |
|
| 42:29 | very prevalent throughout the bacterial world and world. Um And you can use |
|
| 42:35 | number of different terminal acceptor and that's defines an API as something other than |
|
| 42:39 | , as a term acceptor. So can see that we can use um |
|
| 42:44 | nitrate nitrate nitrite generate sulfate. These all options for eco as we're looking |
|
| 42:52 | E Coli here Um and of different donors right form a hydrogen and |
|
| 42:58 | DH lactase. So they can all as donors. So, um so |
|
| 43:04 | combination is used depending on what's available it. Uh But I can certainly |
|
| 43:07 | energy from that. And so it's common for various molecules of nitrogen and |
|
| 43:13 | to be used as electronic sectors. , um uh among, you |
|
| 43:19 | different types of aquatic bacteria and uh bacteria uh can use different oxidized |
|
| 43:26 | of these two elements. Um So let's talk about the uh dissimulator versus |
|
| 43:32 | simulator. So, dissimulator refers to fact that the the the cell is |
|
| 43:37 | on is is not, it's releasing to the environment. So it's dissimulator |
|
| 43:42 | . The the molecule is not held by the cell, it's released to |
|
| 43:45 | environment, right? If it's an simulator process, then the cell is |
|
| 43:50 | on to it. It's gonna use , hold on to it and use |
|
| 43:52 | as an a simulator process. So with using different forms of |
|
| 43:58 | So nitrogen redox a couple. nitrate, nitrate is very popular among |
|
| 44:03 | aerobic Respi respiratory types like E a lot of your uh in tariffs |
|
| 44:09 | use that your E coli, your uh can can uh respiration using nitrate |
|
| 44:17 | reduction of N trite. Um But know, and generally there's not, |
|
| 44:24 | one bacterium has all of these or have a a some of these. |
|
| 44:30 | . Um And what we see, different aspects of, of ni nitrogen |
|
| 44:38 | is the nitrogen cycle. We'll talk that more in, in the chapter |
|
| 44:42 | 22. But uh the nitrogen right, we have um three |
|
| 44:47 | a fixation, nitro application and de . We're in two uh in the |
|
| 44:53 | is, is brought into the either environment or aquatic sediment by a |
|
| 45:00 | OK. And that's in the form ammonia. Ammonia can then be used |
|
| 45:04 | a by lipes can be uh a of electrons. It can be oxidized |
|
| 45:10 | nitrate, nitrite and the nitrate. And of course, that's we call |
|
| 45:16 | is NN, but it's also, a form of lit A as well |
|
| 45:20 | we're using an inorganic electron donor and it to form nitrate and then |
|
| 45:25 | I'm sorry, nit trite can then oxide to form nitrate. Um And |
|
| 45:29 | those are notification reactions. Uh de notification is what we're seeing here. |
|
| 45:35 | this process here that's denitrification. It eventually gets rid of nitrogen from |
|
| 45:46 | system because it gets lost as into ultimately. So that's deification. That's |
|
| 45:51 | form of anaerobic respiration, right? we're using uh either nitrate as a |
|
| 45:56 | acceptor or nit trite or nitric oxide nitrous oxide to nitrogen. So that's |
|
| 46:03 | uh is the gentrification that's of anaerobic represents anaerobic respiration. OK. Um |
|
| 46:14 | , and so I remember it down , this ammonia that's using ammonia as |
|
| 46:23 | electron donor, it becomes oxidized to nitrate. Nitrate gets oxide to form |
|
| 46:30 | . So those are reactions in which the cell can get energy from. |
|
| 46:36 | . The deification is the other end the process. It's terminal acceptor that |
|
| 46:43 | reduced. And so that's anaerobic respiration something other than oxygen, but it |
|
| 46:50 | in the context of nitrogen molecules, represents deification, right, loss of |
|
| 46:54 | from the system. All right, is also uh common especially in marine |
|
| 47:00 | because sulfur is in sulfate uh is present in higher concentrations in marine |
|
| 47:10 | marine ecosystems. So it's very common have sulfate reducing bacteria. Uh or |
|
| 47:15 | uh we're gonna look at a a vent uh environment that would occur in |
|
| 47:22 | uh ocean depths. But here we oxide forms of sulfur, right. |
|
| 47:26 | again, this is respiration using these respiration, using these different forms of |
|
| 47:31 | sulfate to sulfite, sulfide, to sulfate. Uh thos sulfate to sulfur |
|
| 47:37 | , to hydrogen sulfide. These are forms of anaerobic respiration using different forms |
|
| 47:42 | sulfur. Ok. Different oxide forms sulfur. So, when we go |
|
| 47:46 | the depths of the ocean, we see that thermal vents, uh basically |
|
| 47:50 | of it as underwater volcanoes if you . And so they'll, they'll spew |
|
| 47:55 | uh molecules like iron, hydrogen hydrogen gas co2 uh being of |
|
| 48:02 | is pouring out molten lava which is super heat the water So you also |
|
| 48:08 | and generate an environment with different levels , of hypothermic files, thermo Pyles |
|
| 48:14 | meso files. OK? And um tolerance tolerance is the temperature, of |
|
| 48:21 | , as we get farther away from vent, right, that sets of |
|
| 48:25 | where we can have, you your meso, which are have different |
|
| 48:29 | tolerance to heat uh will form farther from the mouth of the vent. |
|
| 48:32 | , but regardless, uh you also of course, different metabolisms, those |
|
| 48:37 | will use things like the sulfide, hydrogen gas and the iron Lit |
|
| 48:46 | using those oxidizing those uh getting right? And they can use the |
|
| 48:51 | to fix CO2, right, little or auto tropes, they fix CO2 |
|
| 48:55 | their carbon. And then of they have the presence of sulfate right |
|
| 49:02 | the oxidation of hydrogen sulfide, right serve as a source for anaerobic respiration |
|
| 49:11 | sulfate reduction in the hydrogen sulfide, example. So these all kind of |
|
| 49:15 | together to, to uh to support other in a way. So products |
|
| 49:21 | one are used as reactants for other . Uh So you have both um |
|
| 49:27 | trophy and a respiration all occurring among different types. Uh So a very |
|
| 49:34 | um um syn trophic, what we a syn trophy working together in terms |
|
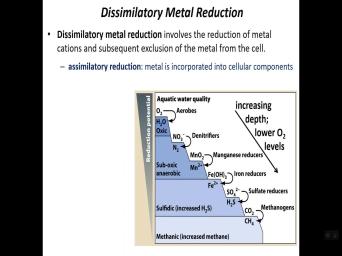
| 49:42 | their metabolism in these environments. So, in dissimulator metal reduction, |
|
| 49:48 | again, this is, this represents respiration. OK. And so you |
|
| 49:54 | and this is what we call an simulator reduction. So the metal ion |
|
| 49:57 | actually incorporated into the central component. it holds on to it, it |
|
| 50:01 | let it go. So that's the . So the simulator metal reduction would |
|
| 50:05 | it gets rid of it. Animator it's going to hold on to it |
|
| 50:08 | use it. OK. And so these kind of what end environments is |
|
| 50:13 | common, our sediments in aquatic environments even in landfills, you can see |
|
| 50:20 | as well. There's um uh landfills be in the top upper layers, |
|
| 50:25 | rich in organic material and that can provide uh fuel for petros aerobic |
|
| 50:31 | So you see here different tiers of uh metabolism. Uh So we have |
|
| 50:37 | potentials which are very positive according to negative reduction potentials. So the upper |
|
| 50:41 | are aerobic, right? You see , OK. Aerobe and oxygen as |
|
| 50:46 | term acceptor. So you have aerobic occurring then you have um and below |
|
| 50:54 | , there would be uh the the fire. So using a nitrate, |
|
| 50:59 | example, uh as a triple So anaerobic respiration then progressively uh processes |
|
| 51:07 | are more and more uh negative in of reduction potential, um especially things |
|
| 51:14 | metal ions, right? Um manganese right are are formed and these can |
|
| 51:21 | more soluble forms of these elements that others can use in the environment. |
|
| 51:25 | it's these are important activities. Uh , to yield these different um molecules |
|
| 51:32 | can be used by others. And you see, the very bottom is |
|
| 51:36 | genesis, this is a process that's easily poisoned by oxygen. So, |
|
| 51:40 | , it's very negative reduction potential. And, and uh typically occurs at |
|
| 51:46 | bottom tier for that reason, it's uh oxygen can uh readily destroy |
|
| 51:51 | process. And so, um uh no, nonetheless, uh methano activity |
|
| 51:56 | methane, which is a actually important gas. So, uh so |
|
| 52:03 | this is representing uh different types of respiration as we go below oxygen, |
|
| 52:09 | course. So your deniro fires, producers, iron reducers, sulfate |
|
| 52:13 | metha are all forms of aerobic And um uh with increasing sensitivities to |
|
| 52:21 | , basically uh more and more negative potentials. So, um the uh |
|
| 52:30 | that basically concludes the uh uh the first, the first part of |
|
| 52:38 | 14, which focuses on electron transport respiration, both aerobic and aerobic. |
|
| 52:43 | you should be familiar with the, comfortable with the concept of of redox |
|
| 52:48 | reduction potential. Uh the strong and donors, strong and weak acceptor. |
|
| 52:53 | so the construction of electronic transport system strong donor to be the acceptor um |
|
| 53:00 | , that's more favorable energetically can produce delta G which it can equate to |
|
| 53:06 | very good proton amount of force. So you should be familiar with, |
|
| 53:10 | that as well, the chemi osmosis . Um And then the, at |
|
| 53:17 | end, we heard we talked about different types of aerobic respiration uh really |
|
| 53:21 | on different nitrogen sulfur molecules as electron and then find the simulator metal |
|
| 53:27 | So, um anyway, so in next uh in part two, |
|
| 53:32 | we'll talk about the photo trophy, li trophy and photo trophy. We'll |
|
| 53:36 | on those two in part two. . Thanks, |
|